Nautilus
| Nautilus Temporal range: Triassic–Present[1] | |
|---|---|
 | |
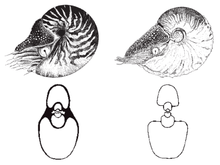
| Nautilus belauensis | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Mollusca |
| Class: | Cephalopoda |
| Subclass: | Nautiloidea |
| Order: | Nautilida |
| Superfamily: | Nautilaceae |
| Family: | Nautilidae Blainville, 1825 |
| Genera | |
|
†Carinonautilus | |
| Synonyms[2] | |
| |
The nautilus (from the Latin form of the original Greek ναυτίλος, 'sailor') is a pelagic marine mollusc of the cephalopod family Nautilidae, the sole extant family of the superfamily Nautilaceae and of its smaller but near equal suborder, Nautilina.
It comprises six living species in two genera, the type of which is the genus Nautilus. Though it more specifically refers to species Nautilus pompilius, the name chambered nautilus is also used for any species of the Nautilidae.
Nautilidae, both extant and extinct, are characterized by involute or more or less convolute shells that are generally smooth, with compressed or depressed whorl sections, straight to sinuous sutures, and a tubular, generally central siphuncle.[3] Having survived relatively unchanged for millions of years, nautiluses represent the only living members of the subclass nautiloidea, and are often considered "living fossils."
The name "nautilus" originally referred to the pelagic octopuses of the genus Argonauta, otherwise known as paper nautiluses, as the ancients believed these animals used their two expanded arms as sails.
Anatomy

Tentacles
Each nautilus tentacle is composed of a long, soft, flexible cirrus and is retractable into a corresponding hardened sheath. Nautiluses typically have more tentacles than other cephalopods — up to ninety. Nautilus tentacles differ from those of other cephalopods. Lacking pads, the tentacles stick to prey by virtue of their ridged surface.[4] Nautiloids have a powerful grip. Attempts to take an object already seized by a nautilus may tear away the creature's tentacles, which will remain firmly attached to the surface of the object.[4] The main tentacles emerge from sheaths which cohere into a single firm fleshy mass. Two pairs of tentacles are separate from the other 90-ish, the pre-ocular and post-ocular, situated before and behind the eye. These are more evidently grooved, with more pronounced ridges. They are extensively ciliated and serve an olfactory purpose.[4][5][6]
Digestive system
The radula is wide and distinctively has nine teeth.
The mouth consists of a parrot-like beak made up of two interlocking jaws capable of ripping the animal's food— mostly crustaceans— from the rocks to which they are attached.[7]:p. 105 Males can be superficially differentiated from females by examining the arrangement of tentacles around the buccal cone: males have a spadix organ (shaped like a spike or shovel) located on the left side of the cone making the cone look irregular, whereas the buccal cone of the female is bilaterally symmetrical.[7]:pp. 115–130
The crop is the largest portion of the digestive tract, and is highly extensible. From the crop, food passes to the small muscular stomach for crushing, and then goes past a digestive caecum before entering the relatively brief intestine.
Circulatory system
Like all cephalopods, the blood of the nautilus contains hemocyanin, which is blue in its oxygenated state. There are two pairs of gills which are the only remnants of the ancestral metamerism to be visible in extant cephalopods.[8]:56 Oxygenated blood arrives at the heart through four ventricles and flows out to the animal's organs through distinct aortas but returns through veins which are too small and varied to be specifically described. The one exception to this is the vena cava, a single large vein running along the underside of the crop into which nearly all other vessels containing de-oxygenated blood empty. All blood passes through one of the four sets of filtering organs (composed of one pericardial appendage and two renal appendages) upon leaving the vena cava and before arriving at the gills for re-oxygenation. Blood waste is emptied through a series of corresponding pores into the pallial cavity.
Nervous system
The central component of the nautilus nervous system is the oesophogeal nerve ring which is a collection of ganglia, commissures, and connectives that together form a ring around the animal's oesophagus. From this ring extend all of the nerves forward to the mouth, tentacles, and funnel; laterally to the eyes and rhinophores; and posteriorly to the remaining organs.
The nerve ring does not constitute what is typically considered a cephalopod "brain": the upper portion of the nerve ring lacks differentiated lobes, and most of the nervous tissue appears to focus on finding and consuming food (i.e., it lacks a "higher learning" center). Nautiluses also tend to have rather short memory spans, and the nerve ring is not protected by any form of brain case.[9]
Shell




Nautiluses are the sole living cephalopods whose bony body structure is externalized as a shell. The animal can withdraw completely into its shell and close the opening with a leathery hood formed from two specially folded tentacles. The shell is coiled, aragonitic,[10] nacreous and pressure resistant, imploding at a depth of about 800 metres (2,600 ft). The nautilus shell is composed of two layers: a matte white outer layer, and a striking white iridescent inner layer. The innermost portion of the shell is a pearlescent blue-gray. The osmeña pearl, contrarily to its name, is not a pearl, but a jewellery product derived from this part of the shell.
Internally, the shell divides into camerae (chambers), the chambered section being called the phragmocone. The divisions are defined by septa, each of which is pierced in the middle by a duct, the siphuncle. As the nautilus matures it creates new, larger camerae, and moves its growing body into the larger space, sealing the vacated chamber with a new septum. The camerae increase in number from around four at the moment of hatching to thirty or more in adults.
The shell colouration also keeps the animal cryptic in the water. When seen from above, the shell is darker in color and marked with irregular stripes, which helps it blend into the dark water below. The underside is almost completely white, making the animal indistinguishable from brighter waters near the surface. This mode of camouflage is named countershading.
The nautilus shell presents one of the finest natural examples of a logarithmic spiral, although it is not a golden spiral. The use of nautilus shells in art and literature is covered at nautilus shell.
Size
Nautilus pompilius is the largest species in the genus. One form from northwestern Australia, once called Nautilus repertus, may reach 26.8 centimetres (10.6 in) in diameter.[11][12] However, most nautilus species never exceed 20 centimetres (7.9 in). Nautilus macromphalus is the smallest species, usually measuring only 16 centimetres (6.3 in). A dwarf population from the Sulu Sea (Nautilus pompilius suluensis) is even smaller, with a mean shell diameter of 115.6 mm.[13]
Physiology
Buoyancy and movement

File format: Ogg
File size: 1.29 MB
Duration: 5.0s

To swim, the nautilus draws water into and out of the living chamber with its hyponome, which uses jet propulsion. While water is inside the chamber, the siphuncle extracts salt from it and diffuses it into the blood. The animal adjusts its buoyancy only in long term density changes by osmosis, either removing liquid from its chambers or allowing water from the blood in the siphuncle to slowly refill the chambers. This is done in response to sudden changes in buoyancy that can occur with predatory attacks of fish, which can break off parts of the shell. This limits nautiluses in that they cannot operate under the extreme hydrostatic pressures found at depths greater than approximately 800 metres (2,600 ft), and in fact implode at about that depth, causing instant death.[13] The gas also contained in the chambers is slightly below atmospheric pressure at sea level.[14] The maximum depth at which they can regulate buoyancy by osmotic removal of chamber liquid is not known.[15]
The nautilus has the extremely rare ability to withstand being brought to the surface from its deep natural habitat without suffering any apparent damage from the experience. Whereas fish or crustaceans brought up from such depths inevitably arrive dead, a nautilus will be unfazed despite the pressure change of as much as 80 atmospheres. The exact reasons for this ability, which is thought to be coincidental rather than specifically functional, are not known, though the perforated structure of the animal's vena cava is thought to play an important role.[7]:p. 188
Senses
.jpg)
Unlike many other cephalopods, nautiluses do not have what many consider to be good vision; their eye structure is highly developed but lacks a solid lens. Whereas a sealed lens allows for the formation of highly focused and clear, detailed surrounding imagery, nautiluses have a simple "pinhole" eye open to the environment which only allows for the creation of correspondingly simple imagery.
Instead of vision, the animal is thought to use olfaction (smell) as the primary sense for foraging and for locating/ identifying potential mates.[16]
The "ear" of the nautilus consists of structures called otocysts located immediately behind the pedal ganglia near the nerve ring. They are oval structures densely packed with elliptical calcium carbonate crystals.
Brain and intelligence
Nautiluses are much closer to the first cephalopods that appeared about 500 million years ago than the early modern cephalopods that appeared maybe 100 million years later (ammonoids and coleoids). They have a seemingly simple brain, not the large complex brains of octopus, cuttlefish and squid, and had long been assumed to lack intelligence. But the cephalopod nervous system is quite different from that of other animals, and recent experiments have shown not only memory, but a changing response to the same event over time.[17][18][19]
In a study in 2008, a group of nautiluses (N. pompilius) were given food as a bright blue light flashed until they began to associate the light with food, extending their tentacles every time the blue light was flashed. The blue light was again flashed without the food 3 minutes, 30 minutes, 1 hour, 6 hours, 12 hours, and 24 hours later. The nautiluses continued to respond excitedly to the blue light for up to 30 minutes after the experiment. An hour later they showed no reaction to the blue light. However, between 6 and 12 hours after the training, they again responded to the blue light, but more tentatively. The researchers concluded that nautiluses had memory capabilities similar to the "short-term" and "long-term memories" of the more advanced cephalopods, despite having different brain structures.[17][18][19] However the long-term memory capability of nautiluses was much shorter than that of other cephalopods. The nautiluses completely forgot the earlier training 24 hours later, in contrast to octopuses, for example, which can remember conditioning for weeks afterwards. However, this may simply be the result of the conditioning procedure being suboptimal for sustaining long-term memories in nautiluses. Nevertheless, the study showed that scientists had previously underestimated the memory capabilities of nautiluses.[19]
Reproduction and lifespan
Nautiluses reproduce by laying eggs. Gravid females attach the fertilized eggs to rocks in shallow waters, whereupon the eggs take eight to twelve months to develop until the 30 millimetres (1.2 in) juveniles hatch. Females spawn once per year and regenerate their gonads, making nautiluses the only cephalopods to present iteroparity or polycyclic spawning.[20]
Nautiluses are sexually dimorphic, in that males have four tentacles modified into an organ, called the "spadix", which transfers sperm into the female's mantle during mating. At sexual maturity, the male shell becomes slightly larger than the female's.[21] Males have been found to greatly outnumber females in practically all published studies, accounting for 60 to 94% of all recorded individuals at different sites.[13]
The lifespan of nautiluses may exceed 20 years, which is exceptionally lengthy for a cephalopod, many of whom live less than three even in captivity and under ideal living conditions.[22] However, nautiluses typically do not reach sexual maturity until they are about 15 years old, limiting their reproductive lifespan to often less than five years.[13]


Right: Shell diameter of mature male and female N. pompilius caught at Osprey Reef. Males (n = 870) had a mean shell diameter of 131.9±2.6 mm, compared to 118.9±7.5 mm in females (n = 86). The Osprey Reef N. pompilius population is the second smallest known in terms of mean shell diameter, after the dwarf form from the Sulu Sea (130.7 mm and 115.6 mm, respectively).[13]
Ecology
Range and habitat

Nautiluses are only found in the Indo-Pacific, from 30° N to 30° S latitude and 90° to 175° E longitude. They inhabit the deep slopes of coral reefs.
Nautiluses usually inhabit depths of several hundred metres. It has long been believed that nautiluses rise at night to feed, mate and lay eggs, but it appears that, in at least some populations, the vertical movement patterns of these animals are far more complex.[23] The greatest depth at which a nautilus has been sighted is 703 m (N. pompilius).[23] Implosion depth for nautilus shells is thought to be around 800 m.[13][23] Only in New Caledonia, the Loyalty Islands, and Vanuatu can nautiluses be observed in very shallow water (at a depth of as little as 5 m).[15][23] This is due to the cooler surface waters found in these southern hemisphere habitats as compared to the many equatorial habitats of other nautilus populations (these usually being restricted to depths greater than 100 m).[15][23] Nautiluses generally avoid water temperatures above 25 °C.[23]

Status
There is growing concern that nautiluses are being greatly overfished not just for the shape of their shells, but also for the nacreous inner shell layer, which is used as a pearl substitute. Because there are currently no national or international regulations protecting this ancient creature, biologist Peter Ward, from the University of Washington, says "there is a horrendous slaughter going on out there."[24] Their limited ecological range and the late onset of their sexual maturity combined with this overfishing has led to recent investigations into the need to protect them from possible endangerment or extinction though no regulations yet exist and thus the nautilus remains unprotected.[25]
In 2016 they were added to CITES II.
Diet
Nautiluses are scavengers and opportunistic predators.[26][27] They eat molts of lobsters, hermit crabs, and carrion of any kind.[15]
Evolution
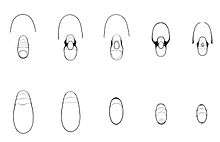
Fossil records indicate that nautiloids have not evolved much during the last 500 million years. Many were initially straight-shelled, as in the extinct genus Lituites. They developed in the Late Cambrian period and became a significant group of sea predators during the Ordovician period. Certain species reached over 2.5 metres (8 ft 2 in) in size. The other cephalopod subclass, Coleoidea, diverged from the nautiloids long ago and the nautilus has remained relatively unchanged since. Nautiloids were much more extensive and varied 200 million years ago. Extinct relatives of the nautilus include ammonites, such as the baculites and goniatites.
The family Nautilidae has its origin in the Trigonocerataceae (Centroceratina), specifically in the Syringonautilidae of the Late Triassic[3] and continues to this day with Nautilus, the type genus, and its close relative, Allonautilus.
Fossil genera
_-_Santiago_specimen.jpg)
The fossil record of Nautilidae begins with Cenoceras in the Late Triassic, a highly varied genus that makes up the Jurassic Cenoceras complex. Cenoceras is evolute to involute, and globular to lentincular; with a suture that generally has a shallow ventral and lateral lobe and a siphuncle that is variable in position but never extremely ventral or dorsal. Cenoceras is not found above the Middle Jurassic and is followed by the Upper Jurassic-Miocene Eutrephoceras.
Eutrephoceras is generally subglobular, broadly rounded laterally and ventrally, with a small to occluded umbilicus, broadly rounded hyponomic sinus, only slightly sinuous sutures, and a small siphuncle that is variable in position.
Next to appear is the Lower Cretaceous Strionautilus from India and the European ex-USSR, named by Shimankiy in 1951. Strionautilus is compressed, involute, with fine longitudinal striations. Whorl sections are subrectangular, sutures sinuous, the siphuncle subcentral.
Also from the Cretaceous is Pseudocenoceras, named by Spath in 1927. Pseudocenoceras is compressed, smooth, with subrectangular whorl sections, flattened venter, and a deep umbilicus. The suture crosses the venter essentially straight and has a broad, shallow, lateral lobe. The siphuncle is small and subcentral. Pseudocenoceras is found in the Crimea and in Libya.
Carinonautilus is a genus from the Upper Cretaceous of India, named by Spengler in 1919. Carinonautilus is a very involute form with high whorl section and flanks that converge on a narrow venter that bears a prominent rounded keel. The umbilicus is small and shallow, the suture only slightly sinuous. The siphuncle is unknown.
Obinautilus has also been placed in Nautilidae by some authorities, though it may instead be an argonautid octopus.[28][29]
Taxonomy

The family Nautilidae contains up to six extant species and several extinct species:
- Genus Allonautilus
- Genus Nautilus
- N. belauensis
- †N. cookanum
- N. macromphalus
- N. pompilius (type)
- N. p. pompilius
- N. p. suluensis
- †N. praepompilius
- N. stenomphalus
Recent genetic data has pointed to there being only three extant species: A. scrobiculatus, N. macromphalus, and N. pompilius, with N. belauensis and N. stenomphalus both subsumed under N. pompilius, possibly as subspecies.[13]
Dubious or uncertain taxa
The following taxa associated with the family Nautilidae are of uncertain taxonomic status:[30]
| Binomial name and author citation | Current systematic status | Type locality | Type repository |
|---|---|---|---|
| N. alumnus Iredale, 1944 | Species dubium [fide Saunders (1987:49)] | Queensland, Australia | Not designated [fide Saunders (1987:49)] |
| N. ambiguus Sowerby, 1848 | Species dubium [fide Saunders (1987:48)] | Not designated | Unresolved |
| N. beccarii Linné, 1758 | Non-cephalopod; Foraminifera [fide Frizzell and Keen (1949:106)] | ||
| N. calcar Linné, 1758 | ?Non-cephalopod; Foraminifera Lenticulina | Adriatic Sea | Unresolved; Linnean Society of London? |
| N. crispus Linné, 1758 | Undetermined | Mediterranean Sea | Unresolved; Linnean Society of London? |
| N. crista Linné, 1758 | Non-cephalopod; Turbo [fide Dodge (1953:14)] | ||
| N. fascia Linné, 1758 | Undetermined | Adriatic Sea | Unresolved; Linnean Society of London? |
| N. granum Linné, 1758 | Undetermined | Mediterranean Sea | Unresolved; Linnean Society of London? |
| N. lacustris Lightfoot, 1786 | Non-cephalopod; Helix [fide Dillwyn (1817:339)] | ||
| N. legumen Linné, 1758 | Undetermined | Adriatic Sea | Unresolved; Linnean Society of London? |
| N. micrombilicatus Joubin, 1888 | Nomen nudum | ||
| N. obliquus Linné, 1758 | Undetermined | Adriatic Sea | Unresolved; Linnean Society of London? |
| N. pompilius marginalis Willey, 1896 | Species dubium [fide Saunders (1987:50)] | New Guinea | Unresolved |
| N. pompilius moretoni Willey, 1896 | Species dubium [fide Saunders (1987:49)] | New Guinea | Unresolved |
| N. pompilius perforatus Willey, 1896 | Species dubium [fide Saunders (1987:49)] | New Guinea | Unresolved |
| N. radicula Linné, 1758 | ?Non-cephalopod; Foraminifera Nodosaria | Adriatic Sea | Unresolved; Linnean Society of London? |
| N. raphanistrum Linné, 1758 | Undetermined | Mediterranean Sea | Unresolved; Linnean Society of London? |
| N. raphanus Linné, 1758 | Undetermined | Adriatic Sea | Unresolved; Linnean Society of London? |
| N. semi-lituus Linné, 1758 | Undetermined | Liburni, Adriatic Sea | Unresolved; Linnean Society of London? |
| N. sipunculus Linné, 1758 | Undetermined | "freto Siculo" | Unresolved; Linnean Society of London? |
| N. texturatus Gould, 1857 | Nomen nudum | ||
| Octopodia nautilus Schneider, 1784 | Rejected specific name [fide Opinion 233, ICZN (1954:278)] |
See also
- Cephalopod size, for maximum shell diameters
- Historiae animalium by Conrad Gessner, first book with fossil illustrations.
- The Nautilus, a malacological journal
- The Chambered Nautilus, a poem of Oliver Wendell Holmes
References
Notes
- ↑ Ward, P. D.; Saunders, W. B. (1997). "Allonautilus: A New Genus of Living Nautiloid Cephalopod and Its Bearing on Phylogeny of the Nautilida". Journal of Paleontology. Paleontological Society. 71 (6): 1054–1064. doi:10.2307/1306604 (inactive 2015-03-04). JSTOR 1306604.
- ↑ Cichowolski, M.; Ambrosio, A.; Concheyro, A. (2005). "Nautilids from the Upper Cretaceous of the James Ross Basin, Antarctic Peninsula". Antarctic Science. 17 (2): 267. doi:10.1017/S0954102005002671.
- 1 2 Kümmel,B. 1964. Nautiloidae-Nautilida, in the Treatise on Invertebrate Paleontology, Geological Society of America and Univ of Kansas Press, Teichert and Moore eds.
- 1 2 3 Willey, Arthur (1897). "The Pre-ocular and Post-ocular Tentacles and Osphradia of Nautilus". Quarterly Journal of Microscopical Science. 40 (1): 197–201.
- ↑ Fukuda, Y. 1987. Histology of the long digital tentacles. In: W.B. Saunders & N.H. Landman (eds.) Nautilus: The Biology and Paleobiology of a Living Fossil. Springer Netherlands. pp. 249–256. doi:10.1007/978-90-481-3299-7_17
- ↑ Kier, W.M. 1987. "The functional morphology of the tentacle musculature of Nautilus pompilius." (PDF). In: W.B. Saunders & N.H. Landman (eds.) Nautilus: The Biology and Paleobiology of a Living Fossil. Springer Netherlands. pp. 257–269. doi:10.1007/978-90-481-3299-7_18
- 1 2 3 Griffin, Lawrence E. (1900). The anatomy of Nautilus pompilius. 8. Washington, D.C.: Government Printing Office. doi:10.5962/bhl.title.10466. OCLC 18760979.
- ↑ Wingstrand, KG (1985). "On the anatomy and relationships of Recent Monoplacophora" (Link to free full text + plates). Galathea Rep. 16: 7–94.
- ↑ Young, J. Z. (27 May 1965). "The Central Nervous System of Nautilus". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. The Royal Society. 249 (754): 1–25. doi:10.1098/rstb.1965.0006. JSTOR 2416631.
- ↑ Buchardt, B.; Weiner, S. (1981). "Diagenesis of aragonite from Upper Cretaceous ammonites: a geochemical case-study". Sedimentology. 28 (3): 423–438. Bibcode:1981Sedim..28..423B. doi:10.1111/j.1365-3091.1981.tb01691.x.
- ↑ Nautilus repertus ID:118764. Shell Encyclopedia, Conchology, Inc.
- ↑ Harasewych, M.G. & F. Moretzsohn (2010). The Book of Shells: A lifesize guide to identifying and classifying six hundred shells. A & C Black Publishers, London.
- 1 2 3 4 5 6 7 8 9 Dunstan AJ; Ward PD; Marshall NJ (February 2011). Solan, Martin, ed. "Nautilus pompilius life history and demographics at the Osprey Reef Seamount, Coral Sea, Australia". PLoS ONE. 6 (2): e16312. Bibcode:2011PLoSO...616312D. doi:10.1371/journal.pone.0016312. PMC 3037366
 . PMID 21347356.
. PMID 21347356. - ↑ Nautilus: The Biology and Paleobiology of a Living Fossil. Springer Netherlands. pp. 552, ch. 34.3.
- 1 2 3 4 Ward, P.D. (1987). The Natural History of Nautilus. Allen and Unwin, London.
- ↑ Grasso, F.; Basil, J. (2009). "The evolution of flexible behavioral repertoires in cephalopod molluscs". Brain, Behavior and Evolution. 74 (3): 231–245. doi:10.1159/000258669. PMID 20029186.
- 1 2 Ewen Callaway (2 June 2008). "Simple-Minded Nautilus Shows Flash of Memory". New Scientist. Retrieved 7 March 2012.
- 1 2 Kathryn Phillips (15 June 2008). "Living Fossil Memories" (PDF). Inside JEB. 211 (12): iii. doi:10.1242/jeb.020370.
- 1 2 3 Robyn Crook & Jennifer Basil (2008). "A biphasic memory curve in the chambered nautilus, Nautilus pompilius L. (Cephalopoda: Nautiloidea)" (PDF). The Journal of Experimental Biology. 211 (12): 1992–1998. doi:10.1242/jeb.018531.
- ↑ Rocha, F.; Guerra, Á.; González, Á. F. (2001). "A review of reproductive strategies in cephalopods". Biological Reviews of the Cambridge Philosophical Society. 76 (3): 291–304. doi:10.1017/S1464793101005681. PMID 11569786.
- ↑ Bruce Saunders, W.; Spinosa, C. (1978). "Sexual Dimorphism in Nautilus from Palau". Paleobiology. 4 (3): 349–358. doi:10.2307/2400210. JSTOR 2400210.
- ↑ Saunders WB (June 1984). "Nautilus Growth and Longevity: Evidence from Marked and Recaptured Animals". Science. 224 (4652): 990–992. Bibcode:1984Sci...224..990S. doi:10.1126/science.224.4652.990. PMID 17731999.
- 1 2 3 4 5 6 Dunstan, A. J.; Ward, P. D.; Marshall, N. J. (2011). Solan, Martin, ed. "Vertical distribution and migration patterns of Nautilus pompilius". PLoS ONE. 6 (2): e16311. doi:10.1371/journal.pone.0016311. PMC 3043052. PMID 21364981.
- ↑ Broad WJ (24 October 2011). "Loving the Chambered Nautilus to Death". New York Times. Retrieved 25 October 2011.
- ↑ Dunstan AJ; Bradshaw CJA; Marshall NJ (February 2011). Solan, Martin, ed. "Nautilus at Risk – Estimating Population Size and Demography of Nautilus pompilius". PLoS ONE. 6 (2): e16716. Bibcode:2011PLoSO...616716D. doi:10.1371/journal.pone.0016716. PMC 3037370. PMID 21347360.
- ↑ Saunders, W.B. (1984). "The role and status of Nautilus in its natural habitat: Evidence from deep-water remote camera photosequences". Paleobiology. 10 (4): 469–486. JSTOR 2400618.
- ↑ Wells, M. J.; Wells, J.; O'Dor, R. K. (2009). "Life at low oxygen tensions: The behaviour and physiology of Nautilus pompilius and the biology of extinct forms". Journal of the Marine Biological Association of the United Kingdom. 72 (2): 313–328. doi:10.1017/S0025315400037723.
- ↑ Teichert, C. & T. Matsumoto (2010). The Ancestry of the Genus Nautilus. In: W.B. Saunders & N.H. Landman (eds.) Nautilus: The Biology and Paleobiology of a Living Fossil. Springer. pp. 25–32. doi:10.1007/978-90-481-3299-7_2
- ↑ Saul, L.R.; Stadum, C.J. (2005). "Fossil argonauts (Mollusca: Cephalopoda: Octopodida) from Late Miocene siltstones of the Los Angeles Basin, California". Journal of Paleontology. 79 (3): 520–531. doi:10.1666/0022-3360(2005)079<0520:FAMCOF>2.0.CO;2.
- ↑ Sweeney, M.J. 2002. Taxa Associated with the Family Nautilidae Blainville, 1825. Tree of Life web project.
Bibliography
- Ward, P.D. 1988. In Search of Nautilus. Simon and Schuster.
- W. Bruce Saunders & Neil H. Landman (2010). Nautilus: the biology and palaeontology of a living fossil. Topics in Geobiology. 6. Dordrecht : Springer Science+Business Media B.V. ISBN 978-90-481-3299-7.
External links
| Wikimedia Commons has media related to Nautilus. |
- Nautilidae discussion forum, tonmo.com
- Waikïkï Aquarium: Marine Life Profile: Chambered Nautilus, waguarium.org
- A molecular and karyological approach to the taxonomy of Nautilus, utmb.edu