Borrelia burgdorferi
| Borrelia burgdorferi | |
|---|---|
_lores.jpg) | |
| Borrelia burgdorferi | |
| Scientific classification | |
| Domain: | Bacteria |
| Phylum: | Spirochaetes |
| Order: | Spirochaetales |
| Family: | Spirochaetaceae |
| Genus: | Borrelia |
| Species: | B. burgdorferi |
| Binomial name | |
| Borrelia burgdorferi Johnson et al. 1984 emend. Baranton et al. 1992 | |
Borrelia burgdorferi is a bacterial species of the spirochete class of the genus Borrelia. B. burgdorferi exists in North America and Europe and is the predominant causative agent of Lyme disease in the United States. Borrelia species are considered diderm (double-membrane) bacteria rather than Gram-positive or negative.[1]
Microbiology
Borrelia burgdorferi is named after the researcher Willy Burgdorfer, who first isolated the bacterium in 1982.[2]
Morphology
B. burgdorferi resembles other spirochetes in that it has an outer membrane and inner membrane with a thin layer of peptidoglycan in between. However, the outer membrane lacks lipopolysaccharide. Its shape is a flat wave. It is about 0.3 μm wide and 5 to 20 μm in length.[3]
B. burgdorferi is an anaerobic, motile spirochete with seven to 11 bundled perisplasmic flagella set at each end that allow the bacterium to move in low- and high-viscosity media alike, which is related to its high virulence factor.[4]
Metabolism
B. burgdorferi is a slow-growing microaerophilic spirochete with a doubling time of 24 to 48 hours.[5] It is one of the few bacteria that can survive without iron, having replaced all of its iron-sulfur cluster enzymes with enzymes that use manganese, thus avoiding the problem many pathogenic bacteria face in acquiring iron.[6]
Life cycle
B. burgdorferi circulates between Ixodes ticks and a vertebrate host in an enzootic cycle. B. burgdorferi living in a tick cannot be passed to its offspring. Therefore, ticks must feed on the blood of an infected vertebrate to acquire B. burgdorferi. Infected ticks transmit B. burgdorferi by feeding on another vertebrate to complete the cycle.[7] Ticks can transmit B. burgdorferi to humans, but humans are dead-end hosts, unlikely to continue the life cycle of the spirochete.[8]
Disease
Lyme disease is a zoonotic, vector-borne disease transmitted by the Ixodes tick (also the vector for Babesia). The infected nymphal tick transmits B. burgdorferi via its saliva to the human during its blood meal.[9] In order for a successful infection, the vertebrate host reservoir must cultivate enough bacteria that can be circulated throughout the blood, so that B. burgodorferi can be transmitted through Ixodes blood feeding.[7] Additionally, the bacteria itself must withstand the molting and life cycle of the Ixodes tick and successfully transinfect a host for B. Burgdorferi to spread to humans.[7]
Clinical presentation of Lyme disease may include the characteristic bull's-eye rash and erythema chronicum migrans (a rash which spreads peripherally and spares the central part), as well as myocarditis, cardiomyopathy, arrythmia, arthritis, arthralgia, meningitis, neuropathies, and facial nerve palsy.[10]
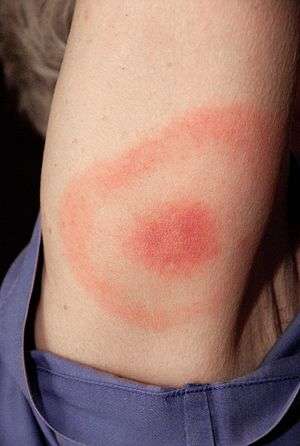
B. burgdorferi infections have been found in possible association with primary cutaneous B-cell lymphomas (PCBCLs),[11][12] where a review of the primary literature has, as of 2010, noted that most of the PCBLCs examined have been 'unresponsive' to antibiotics;[12]:846 hence, as in case of Chlamydophila psittaci association with ocular adnexal mucosa-associated lymphoid tissue (MALT) lymphoma, the working conclusion was that "if B. burgdorferi is truly associated with PCBCL, then there is wide geographic variability and other factors are probably involved".[12]:846
Progression of the disease follows from 3 stages.
Stage 1
Stage 1 affects the area around the bite, with a rash or swelling possible.
Stage 2
Stage 2 occurs weeks to months after; if left untreated, the bacteria spread through the body and affect the heart, bones, and nervous system.
Stage 3
Stage 3 occurs years after and chronic arthritis and neurological complications develop.[10]
Anaplasmosis and babesiosis are also common tick borne pathogens that infect humans similarly to Borrelia burgdorferi.[13] Consequently, it is possible for an Ixodes tick to coinfect a host with either two or all other diseases. When a host is coinfected, the combined effects of the diseases act synergistically, often proving to cause worse symptoms than a single infection alone[13] Coinfected humans tend to present with a more severe manifestation of Lyme disease. In addition, they tend to acquire a wider range of secondary symptoms, such as influenza-like symptoms.[13] More studies and research must be done to determine the synergistic effect of co-infection and its effect on the human body.
Variation of Severity
So far, there are three factors that may contribute to the severity of the clinical manifestation of Lyme Disease. The presence of ribosomal spacers, plasmids, and the outer surface protein C (OspC) are indicators of the severity of the infection.[14] Additionally, humans, themselves, vary in their response to the infection.[14] The variation in response leads to different clinical manifestations and different infections to different organs.
Molecular pathogenesis
After the pathogen is transmitted, it will acclimate to the mammalian conditions. Borrelia burgdorferi will change its glycoproteins and proteases on its plasma membrane to facilitate its dissemination throughout the blood.[14] While infecting, B. burgdorferi will express proteins that will interact with endothelial cells, platelets, chondrocytes, and the extracellular matrix.[14] This interaction inhibits proper function of the infected areas, leading to the pathological manifestations of Lyme disease. In response, the host will initiate an inflammatory response to attempt to remove the infection.[14]
Borrelia burgdorferi, also, expresses at least seven plasminogen binding proteins for interference of factor H at the activation level. This is part of a complement system evasion strategy that leads to downstream blocking of immune response.[15]
Genetics
B. burgdorferi (B31 strain) was the third microbial genome ever sequenced, following the sequencing of both Haemophilus influenzae and Mycoplasma genitalium in 1995. Its linear chromosome contains 910,725 base pairs and 853 genes.[16] The sequencing method used was whole genome shotgun. The sequencing project, published in Nature in 1997 and Molecular Microbiology in 2000, was conducted at The Institute for Genomic Research.[17] Overall, B. burgdorferi's genome oddly consists of one megabase chromosome and a variety of circular and linear plasmids ranging in size from 9 to 62 kilobases.[7] The megabase chromosome, unlike many other eubacteria, has no relation to either the bacteria's virulence or to the host-parasite interaction.[16] Some of the plasmids are necessary for the B. burgdorferi life cycle but not for propagation of the bacteria in culture.[7]
The genomic variations of Borrelia burgdorferi contribute to varying degrees of infection and dissemination.[18] Each genomic group has varying antigens on its membrane receptor, which are specific to the infection of the host. One such membrane receptor is the surface protein OspC.[18] The OspC surface protein is shown to be a strong indicator of the identification of genomic classification and the degree of dissemination.[18] Varying number of OspC loci are indications and determinants for the variations of Borrelia burgdorferi.[18] The surface protein is also on the forefront of current vaccine research for Lyme disease via Borrelia.[19]
Evolution
Genetically diverse B. burgdorferi strains, as defined by the sequence of ospC, are maintained within the Northeastern United States. Balancing selection may act upon ospC or a nearby sequence to maintain the genetic variety of B. burgdorferi.[20] Balancing selection is the process by which multiple versions of a gene are kept within the gene pool at unexpectedly high frequencies. Two major models that control the selection balance of B.burgdorferi is negative frequency-dependent selection and multiple-niche polymorphism[21]. These models may explain how B. burgdorferi have diversified, and how selection may have affected the distribution of the B. burgdorferi variants, or the variation of specific traits of the species, in certain environments.
Negative-Frequency Dependent Selection
In negative frequency-dependent selection, rare and uncommon variants will have a selective advantage over variants that are very common in an environment.[21] For B. burgdorferi, low-frequency variants will be advantageous because potential hosts will not have a built immunological response to the variants specific OspC outer protein.[21]
Multiple-niche Polymorphism
Ecological niches are all of the variables in an environment, such as the resources, competitors, and responses, that contribute to the organism's fitness. Multiple-niche polymorphism states that diversity is maintained within a population due to the varying amount of possible niches and environments.[21] Therefore, the more various niches the more likelihood of polymophrism and diversity. For B. burgdorferi, varying vertebrae niches, such deer and mice, can affect the overall balancing selection for variants.[21]
See also
Further reading
- Velázquez, Encarna, Peix, Álvaro & Gómez-Alonso, Alberto, 2011, "Microorganismos y cáncer: evidencias científicas y nuevas hipótesis", Cirugía Española, vol 89, no. 3, pages 136–144. ISSN 0009-739X; doi 10.1016/j.ciresp.2010.08.006; accessed 16 July 2015. English translation
References
- ↑ Samuels DS; Radolf, JD, eds. (2010). "Chapter 6, Structure, Function and Biogenesis of the Borrelia Cell Envelope". Borrelia: Molecular Biology, Host Interaction and Pathogenesis. Caister Academic Press. ISBN 978-1-904455-58-5.
- ↑ Burgdorfer W, Barbour AG, Hayes SF, Benach JL, Grunwaldt E, Davis JP (June 1982). "Lyme disease-a tick-borne spirochetosis?". Science. 216 (4552): 1317–9. Bibcode:1982Sci...216.1317B. doi:10.1126/science.7043737. PMID 7043737.
- ↑ Motaleb MA, Liu J, Wooten RM (2015). "Spirochetal motility and chemotaxis in the natural enzootic cycle and development of Lyme disease". Current Opinion in Microbiology. 28: 106–13. doi:10.1016/j.mib.2015.09.006. PMC 4688064
 . PMID 26519910.
. PMID 26519910. - ↑ Motaleb, Mohammed; Corum, Linda; Bono, James; Elias, Abdallah; Rosa, Patricia; Samuels, D. Scott; Charon, Nyles (2000). "Borrelia burgdorferi periplasmic flagella have both skeletal and motility functions". Proceedings of the National Academy of Sciences of the United States of America. 97 (20): 10899–10904. Bibcode:2000PNAS...9710899M. doi:10.1073/pnas.200221797. PMC 27121. PMID 10995478.
- ↑ Zückert WR (2007). "Laboratory maintenance of Borrelia burgdorferi". Current Protocols in Microbiology. Chapter 12: Unit 12C.1. doi:10.1002/9780471729259.mc12c01s4. ISBN 0471729256. PMID 18770608.
- ↑ Galdwin, Mark; Trattler, Bill (2009). Spirochetes: Clinical Microbiology Made Ridiculously Simple. MedMaster, Inc. ISBN 978-0-940780-81-1.
- 1 2 3 4 5 Brisson D, Drecktrah D, Eggers CH, Samuels DS (2012). "Genetics of Borrelia burgdorferi". Annual Review of Genetics. 46: 515–36. doi:10.1146/annurev-genet-011112-112140. PMC 3856702. PMID 22974303.
- ↑ Radolf JD, Caimano MJ, Stevenson B, Hu LT (2012). "Of ticks, mice and men: understanding the dual-host lifestyle of Lyme disease spirochaetes". Nature Reviews. Microbiology. 10 (2): 87–99. doi:10.1038/nrmicro2714. PMC 3313462. PMID 22230951.
- ↑ Tilly K, Rosa PA, Stewart PE (2008). "Biology of infection with Borrelia burgdorferi". Infectious Disease Clinics of North America. 22 (2): 217–34, v. doi:10.1016/j.idc.2007.12.013. PMC 2440571. PMID 18452798.
- 1 2 "Signs and Symptoms, Lyme Disease". Centers For Disease Control. March 4, 2015. Retrieved 2015-07-16.
- ↑ Guidoboni M, Ferreri AJ, Ponzoni M, Doglioni C, Dolcetti R (January 2006). "Infectious agents in mucosa-associated lymphoid tissue-type lymphomas: pathogenic role and therapeutic perspectives". Clinical Lymphoma & Myeloma. 6 (4): 289–300. doi:10.3816/CLM.2006.n.003. PMID 16507206.
- 1 2 3 Chang, A. H.; Parsonnet, J. (2010). "Role of Bacteria in Oncogenesis" (PDF). Clinical Microbiology Reviews. 23 (4): 837–857. doi:10.1128/CMR.00012-10. ISSN 0893-8512. PMC 2952975. PMID 20930075.
- 1 2 3 Swanson, Stephen J.; Neitzel, David; Reed, Kurt D.; Belongia, Edward A. (2006-10-01). "Coinfections Acquired from Ixodes Ticks". Clinical Microbiology Reviews. 19 (4): 708–727. doi:10.1128/CMR.00011-06. ISSN 0893-8512. PMC 1592693. PMID 17041141.
- 1 2 3 4 5 Weis, Janet (2011). "Critical Needs and Gaps in Understanding Prevention, Amelioration, and Resolution of Lyme and Other Tick-Borne Diseases: The Short-Term and Long-Term Outcomes: Workshop Report.". The National Academies: 97–101.
- ↑ Zipfel, P., Hallström, T., & Riesbeck, K. (2013). Human complement control and complement evasion by pathogenic microbes – Tipping the balance. Molecular Immunology, 56(3), 152-160.
- 1 2 Fraser CM, Casjens S, Huang WM, et al. (December 1997). "Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi". Nature. 390 (6660): 580–6. Bibcode:1997Natur.390..580F. doi:10.1038/37551. PMID 9403685.
- ↑ Casjens S, Palmer N, van Vugt R, Huang WM, Stevenson B, Rosa P, Lathigra R, Sutton G, Peterson J, Dodson RJ, Haft D, Hickey E, Gwinn M, White O, Fraser CM (2000). "A bacterial genome in flux: the twelve linear and nine circular extrachromosomal DNAs in an infectious isolate of the Lyme disease spirochete Borrelia burgdorferi". Molecular Microbiology. 35 (3): 490–516. doi:10.1046/j.1365-2958.2000.01698.x. PMID 10672174.
- 1 2 3 4 Theisen, M.; Borre, M.; Mathiesen, M. J.; Mikkelsen, B.; Lebech, A. M.; Hansen, K. (1995-06-01). "Evolution of the Borrelia burgdorferi outer surface protein OspC.". Journal of Bacteriology. 177 (11): 3036–3044. ISSN 0021-9193. PMC 176990. PMID 7768799.
- ↑ Embers, Monica E.; Narasimhan, Sukanya (2013-02-12). "Vaccination against Lyme disease: past, present, and future". Frontiers in Cellular and Infection Microbiology. 3: 6. doi:10.3389/fcimb.2013.00006. ISSN 2235-2988. PMC 3569838. PMID 23407755.
- ↑ Kurtenbach K, Hanincová K, Tsao JI, Margos G, Fish D, Ogden NH (2006). "Fundamental processes in the evolutionary ecology of Lyme borreliosis". Nature Reviews. Microbiology. 4 (9): 660–9. doi:10.1038/nrmicro1475. PMID 16894341.
- 1 2 3 4 5 Samuels, D. Scott (2010-01-01). Borrelia: Molecular Biology, Host Interaction and Pathogenesis. Horizon Scientific Press. ISBN 9781904455585.
External links
| Wikispecies has information related to: Borrelia burgdorferi |
- Video Interview, Eva Sapi, PhD on Borrelia Biofilms
- NCBI Borrelia Taxonomy Browser
- Borrelia burgdoferi B31 Genome Page