Serine hydroxymethyltransferase
| Serine Hydroxymethyltransferase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 PyMol rendered crystal structure of serine hydroxymethyltransferase | |||||||||
| Identifiers | |||||||||
| EC number | 2.1.2.1 | ||||||||
| CAS number | 9029-83-8 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Serine hydroxymethyltransferase (SHMT) is a PLP dependent enzyme (EC 2.1.2.1) which plays an important role in cellular one-carbon pathways by catalyzing the reversible, simultaneous conversions of L-serine to glycine and tetrahydrofolate (THF) to 5,10-methylenetetrahydrofolate.[1] This reaction provides the largest part of the one-carbon units available to the cell.[2]
Structure
The structure of the SHMT monomer is remarkably similar across prokaryotes and eukaryotes, but whereas the active enzyme in solution is a dimer in prokaryotes, the enzyme exists as a tetramer in eukaryotic cells, though the evolutionary basis for this difference in structure is not clear.[1] However, the evolutionary path taken by SHMT going from prokaryotic dimeric form to the eukaryotic tetrameric form can be easily seen as a sort of doubling event. In other words, the eukaryotic SHMT tetramer resembles two prokaryotic dimers that have packed together, forming what has been described as a “dimer of dimers.”[3] The interaction between two monomers within a dimer subunit has been found to occur over a greater contact area and is thus much tighter than the interaction between the two dimers.[3]
A single SHMT monomer can be understood in terms of three domains: an N-terminus “arm,” a “large” domain, and a “small” domain.[3] The N-terminus arm appears to be critical for the maintenance of the tight interaction between two monomers. The arm, consisting of two alpha helices and a beta sheet, wraps around the other monomer when in oligomeric form.[3] The “large” domain contains the PLP binding site, which resembles the PLP binding domains of related PLP dependent proteins, like aspartate aminotransferase.[3] The large domain in the eukaryotic form also contains a histidine that has been found essential for tetramer stability.[3] All four histidines of these residues, one from each monomer, sit at the center of the tetrameric complex, where two histidines from a dimeric subunit engages in stacking interactions with the histidines of the other subunit.[3] Prokaryotic SHMT has a proline residue rather than histidine in the equivalent position, which would in part explain why prokaryotic SHMT does not form tetramers.[4]
The active site structure is also quite conserved across eukaryotic and prokaryotic forms. The PLP is anchored by means of a lysine, which forms an aldimine Schiff base linkage with the PLP aldehyde.[5] It has been hypothesized that a nearby tyrosine acts as the proton donor and acceptor during the transaldimination step as well as the formyl transfer step and that an arginine residue engages the tyrosine side chain in a cation-π interaction, which helps to lower the pKa of the tyrosine, making proton transfers more facile.[5]
Mechanism
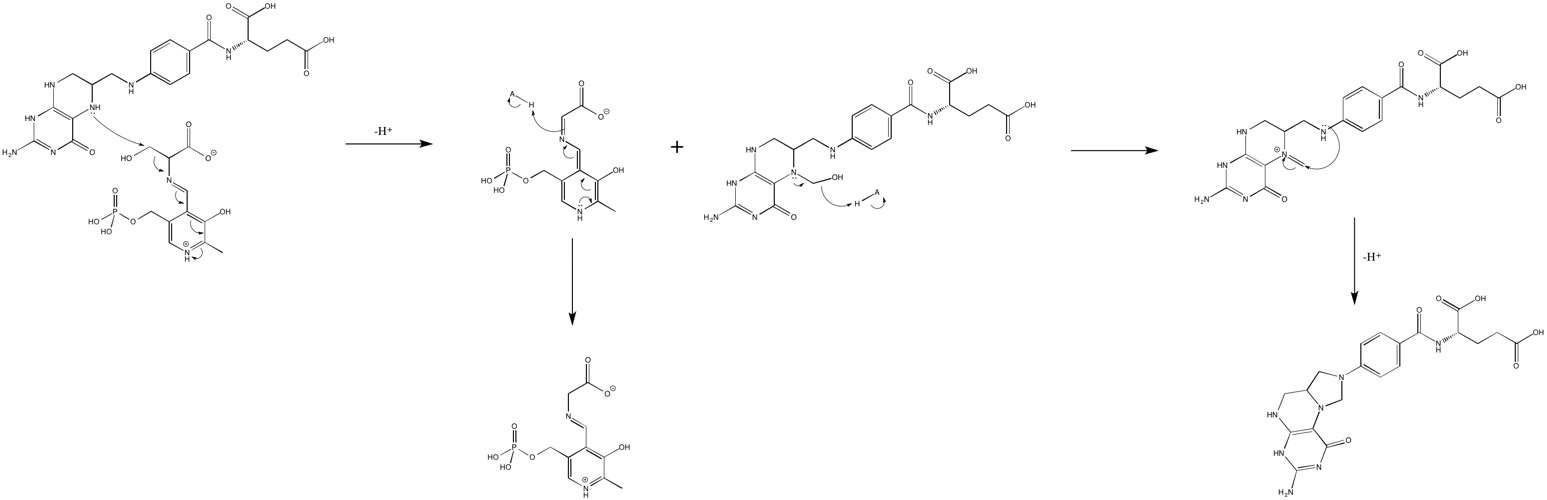
The mechanism commonly ascribed to SHMT enzymatic activity is a transamidation followed by a cleavage of amino acid side chain from the backbone.[5] The N-terminal amine of serine makes a nucleophilic attack on the aldimine between the SHMT lysine and the PLP aldehyde to form a gem-diamine, and then the N-terminal amine lone pair comes down to kick out the lysine, forming a new aldimine, this time with the serine.[5][6] It is believed that a nearby tyrosine is responsible for much of the proton transfers that occur during the transaldimination.[5]
Once the serine is bound to PLP, the nucleophilic amines on THF then attacks the free formaldehyde to make the carbinolamine intermediate.[6] In the second case, the nucleophilic amines on THF attack the serine side chain carbon, simultaneously forming a carbinolamine intermediate on the THF and a quinoid intermediate with the PLP.[6][7] However, THF is not an obligate substrate for SHMT, meaning the cleavage of serine and other ß-hydroxy amino acids (Like threonine) can occur without the presence of THF, and in this case, the mechanism is a retro-aldol cleavage.[8] Also, it seems that the subsequent dehydration of the carbinolamine intermediate to form the methylene bridge and fully cyclize into 5, 10-CH2-THF is not catalyzed by the enzyme, and this reaction may just occur spontaneously.[6]
Clinical Significance
Folate metabolism has already been the subject of chemotherapeutic strategies, but SHMT inhibition, while researched, had not really been taken advantage of in commercial anticancer drugs.[9] However, because the folates used by folate metabolic and folate dependent enzymes are all very similar in structure, and folate mimics being already common in medical use, it has not been difficult to find potential molecular structures that may inhibit SHMT.[9] For example, pemetrexed is already used as an antifolate to treat mesothelioma and was found to be an effective inhibitor of SHMT,[9] and screening other antifolates revealed lometrexol as another effective inhibitor of SHMT.[10]
SHMT has also undergone investigation as a potential target for antimalarial drugs. Research indicates that the active site environment of Plasmodium SHMTs (PSHMTs) differ from that of human cytosolic SHMT quite a bit, allowing for the possibility of selective inhibition of PSHMT and, thus, the treatment of malaria infections.[11] In particular, certain pyrazolopyran molecules have been shown to have a selective nanomolar efficacy against PSHMTs. Poor pharmacokinetics, however, have prevented these pyrazolopyrans from being effective in living models.[12]
Isoforms
Bacteria such as Escherichia coli and Bacillus stearothermophilus have versions of this enzyme and there appear to be two isoforms of SHMT in mammals, one in the cytoplasm (cSHMT) and another in the mitochondria (mSHMT).[1] Plants may have an additional SHMT isoform within chloroplasts.[13]
In mammals, the enzyme is a tetramer of four identical subunits of approximately 50,000 Daltons each. The intact holoenzyme has a molecular weight of approximately 200,000 Daltons and incorporates four molecules of pyridoxal phosphate (Vitamin B6) as a coenzyme.[14]
Other reactions
As well as its primary role in folate metabolism, SHMT also catalyzes other reactions that may be biologically significant, including the conversion of 5,10-methenyltetrahydrofolate to 10-formyltetrahydrofolate.[2] When coupled with C1-tetrahydrofolate synthase and tetrahydropteroate, cSHMT also catalyzes the conversion of formate to serine.[2]
Role in Smith-Magenis syndrome
Smith-Magenis syndrome (SMS) is a rare disorder that manifests as a complex set of traits including facial abnormalities, unusual behaviors, and developmental delay.[15] It results from an interstital deletion within chromosome 17p11.2, including the cSHMT gene and a small study showed SHMT activity in SMS patients was ~50% of normal.[15] Reduced SHMT would result in less glycine which could affect the nervous system by acting as an agonist to the NMDA receptor and this could be a mechanism behind SMS.[15]
References
- 1 2 3 Appaji Rao N, Ambili M, Jala VR, Subramanya HS, Savithri HS (April 2003). "Structure-function relationship in serine hydroxymethyltransferase". Biochim. Biophys. Acta. 1647 (1–2): 24–9. doi:10.1016/s1570-9639(03)00043-8. PMID 12686103.
- 1 2 3 Stover P, Schirch V (August 1990). "Serine hydroxymethyltransferase catalyzes the hydrolysis of 5,10-methenyltetrahydrofolate to 5-formyltetrahydrofolate". J. Biol. Chem. 265 (24): 14227–33. PMID 2201683.
- 1 2 3 4 5 6 7 Renwick S, Snell K, Baumann U (1998). "The crystal structure of human cytosolic serine hydroxymethyltransferase: a target for cancer chemotherapy.". Structure. 6 (9): 1105–1116. doi:10.1016/s0969-2126(98)00112-9.
- ↑ Scarsdale J, Radaev S, Kazanina G, Schirch V, Wright, H (2000). "Crystal structure at 2.4 A resolution of E. coli serine hydroxymethyltransferase in complex with glycine substrate and 5-formyl tetrahydrofolate.". J. Mol. Bio. 296 (1): 155–168. doi:10.1006/jmbi.1999.3453.
- 1 2 3 4 5 Florio R, di Salvo M, Vivoli M, Contestabile R (2011). "erine hydroxymethyltransferase: a model enzyme for mechanistic, structural, and evolutionary studies.". Biochim. Biophys. Acta. 1814 (11): 1489–1496. doi:10.1016/j.bbapap.2010.10.010.
- 1 2 3 4 Schirch V, Szebenyi D (2005). "Serine hydroxymethyltransferase revisited.". Curr. Opin. Chem. Bio. 9 (5): 482–487. doi:10.1016/j.cbpa.2005.08.017.
- ↑ Szebenyi D, Musayev F, di Salvo M, Safo M, Schirch V (2004). "Serine hydroxymethyltransferase: role of glu75 and evidence that serine is cleaved by a retro-aldol mechanism.". Biochemistry. 43 (22): 6865–6876. doi:10.1021/bi049791y.
- ↑ Chiba Y, Terada T, Kameya M, Shimizu K, Arai H, Ishii M, Igarashi Y (2012). "Mechanism for folate-independent aldolase reaction catalyzed by serine hydroxymethyltransferase.". FEBS. 279 (3): 504–514. doi:10.1111/j.1742-4658.2011.08443.x.
- 1 2 3 Daidone F, Florio R, Rinaldo S, Contestabile R, di Salvo M, Cutruzzola F, Bossa F, Paiardini A (2011). "In silico and in vitro validation of serine hydroxymethyltransferase as a chemotherapeutic target of the antifolate drug pemetrexed.". Eur. J. Med. Chem. 46 (5): 1616–1621. doi:10.1016/j.ejmech.2011.02.009.
- ↑ Paiardini A, Fiascarelli A, Rinaldo S, Daidone F, Giardina G, Koes D, Parroni A, Montini G, Marani M, Paone A, McDermott L, Contestabile R, Cutruzzola F (2015). "Screening and in vitro testing of antifolate inhibitors of human cytosolic serine hydroxymethyltransferase.". Chem. Med. Chem. 10 (3): 490–497. doi:10.1002/cmdc.201500028.
- ↑ Pinthong C, Maenpuen S, Amornwatcharapong W, Yuthavong Y, Leartsakulpanich U, Chaiyen P (2014). "Distinct biochemical properties of human serine hydroxymethyltransferase compared with the Plasmodium enzyme: implications for selective inhibition.". FEBS. 281 (11): 2570–2583. doi:10.1111/febs.12803.
- ↑ Witschel M, Rottmann M, Schwab A, Leartsakulpanich U, Chitnumsub P, Seet M, Tonazzi S, Schwertz G, Stelzer F, Mietzner T, McNamara C, Thater F, Freymond C, Jaruwat A, Pinthong C, Riangrungroj P, Oufir M, Hamburger M, Maser P, Sanz-Alonso L, Charman S, Wittlin S, Yuthavong Y, Chaiyen P, Diederich F (2015). "Inhibitors of plasmodial serine hydroxymethyltransferase (SHMT): cocrystal structures of pyrazolopyrans with potent blood- and liver-stage activities.". J. Med. Chem. 58 (7): 3117–3130. doi:10.1021/jm501987h.
- ↑ Besson V, Nauburger M, Rebeille F, Douce R (1995). "Evidence for three serine hydroxymethyltransferases in green leaf cells. Purification and characterization of the mitochondrial and chloroplastic isoforms". Plant Physiol. Biochem. 33 (6): 665–673.
- ↑ Martinez-Carrion M, Critz W, Quashnock J (April 1972). "Molecular weight and subunits of serine transhydroxymethylase". Biochemistry. 11 (9): 1613–5. doi:10.1021/bi00759a011. PMID 5028104.
- 1 2 3 Elsea SH, Juyal RC, Jiralerspong S, Finucane BM, Pandolfo M, Greenberg F, Baldini A, Stover P, Patel PI (December 1995). "Haploinsufficiency of cytosolic serine hydroxymethyltransferase in the Smith-Magenis syndrome". Am. J. Hum. Genet. 57 (6): 1342–50. PMC 1801426
 . PMID 8533763.
. PMID 8533763.
Literature
- Akhtar M, el-Obeid HA (1972). "Inactivation of serine transhydroxymethylase and threonine aldolase activities". Biochim. Biophys. Acta. 258 (3): 791–9. doi:10.1016/0005-2744(72)90180-5. PMID 5017703.
- Blakley RL (1960). "A spectrophotometric study of the reaction catalysed by serine transhydroxymethylase". Biochem. J. 77 (3): 459–65. PMC 1205057. PMID 16748851.
- Fujioka M (1969). "Purification and properties of serine hydroxymethylase from soluble and mitochondrial fractions of rabbit liver". Biochim. Biophys. Acta. 185 (2): 338–49. doi:10.1016/0005-2744(69)90427-6. PMID 5808700.
- Kumagai H, Nagate T, Yoshida H, Yamada H (1972). "Threonine aldolase from Candida humicola. II. Purification, crystallization and properties". Biochim. Biophys. Acta. 258 (3): 779–90. doi:10.1016/0005-2744(72)90179-9. PMID 5017702.
- Schirch L, Gross T (1968). "Serine transhydroxymethylase. Identification as the threonine and allothreonine aldolases". J. Biol. Chem. 243 (21): 5651–5. PMID 5699057.
- Schirch L, Quashnock, J (1981). "Evidence that tetrahydrofolate does not bind to serine hydroxymethyltransferase with positive homotropic cooperativity". J. Biol. Chem. 256 (12): 6245–9. PMID 6787050.
- Quashnock J, Chlewboski J, Martinez-Carrion M, Schirch L (1983). "Serine Hydroxymethyltransferase 31P Nuclear Magnetic Resonance Study of the Enzyme-Bound Pyridoxal 5'-Phosphate". J. Biol. Chem. 258 (1): 503–7. PMID 6848517.
External links
- Serine Hydroxymethyltransferase at the US National Library of Medicine Medical Subject Headings (MeSH)