Pursuit predation
Pursuit predation is a form of predation in which predators give chase to fleeing prey. The chase can be initiated either by the predator or by the prey, should the prey be alerted to a predator's presence and attempt to flee before the predator gives chase. The chase ends with either the predator capturing and consuming the prey, effectively diminishing the prey's fitness, or with the prey escaping the predator's hunt, thus maintaining the prey's overall fitness, but leaving both prey and predator with metabolic losses. Pursuit predation is typically observed in carnivorous species within the kingdom Animalia, with some iconic examples being cheetahs, lions, and wolves.
Pursuit predation is an alternate predation strategy to ambush predation. While pursuit predators use a detection and pursuit phase in order to obtain prey, ambush predators use stealth to capture prey. Strength and speed are important to pursuit predators, whereas ambush predators ignore these in favor of surprise from a typically concealed location. While the two patterns of predation are not mutually exclusive, morphological differences in body plan can create a bias in an organism towards each type of predation.
Strategy
There is still uncertainty as to whether predators behave with a general tactic or strategy while preying.[1] However, among pursuit predators, there are several common behaviors. Often, predators will scout potential prey, assessing prey quantity and density prior to engaging in a pursuit. Certain predators choose to pursue prey primarily in a group of conspecifics; such animals are known as pack hunters or group pursuers. Other species choose to hunt alone. These two behaviors are typically due to differences in hunting success, where some groups are very successful in groups and others are more successful alone. Pursuit predators may also choose to either exhaust their metabolic resources rapidly [2] or pace themselves during a chase.[3] This choice can influenced by prey species, seasonal settings, or temporal settings. Predators that rapidly exhaust their metabolic resources during a chase tend to first stalk their prey, slowly approaching their prey to decrease chase distance and time. When the predator is at a closer distance (one that would lead to easier prey capture), it finally gives chase.[4] Pacing pursuit is more commonly seen in group pursuit, as individual animals do not need to exert as much energy to capture prey. However, this type of pursuit requires group coordination, which may have varying degrees of success. Since groups can engage in longer chases, they often focus on separating a weaker or slower prey item during pursuit.[5] Morphologically speaking, while ambush predation requires stealth,[6] pursuit predation requires speed; pursuit predators are proportionally long-limbed and equipped with cursorial adaptations.[7] Current theories suggest that this proportionally long-limbed approach to body plan was an evolutionary countermeasure to prey adaptation.[7]
Group Pursuers
Vertebrates

Group pursuers hunt with a collection of conspecifics. Group pursuit is usually seen in species of relatively high sociality; in vertebrates, individuals often seem to have defined roles in pursuit.
Mammals
African wild dog (Lycaon pictus) packs have been known to split into several smaller groups while in pursuit; one group initiates the chase, while the other travels ahead of the prey's escape path. The group of chase initiators coordinate their chase to lead the prey towards the location of the second group, where the prey's escape path will be effectively cut off.[8] Bottlenose dolphins (Tursiops) have been shown exhibiting similar behaviors of pursuit role specialization. One group within the dolphin pod, known as the drivers, give chase to the fish - forcing the fish into a tight circle formation, while the other group of the pod, the barriers, approach the fish from the opposite direction. This two-pronged attack leaves the fish with only the option of jumping out of the water to escape the dolphins. However, the fish are completely vulnerable in the air; it is at this point when the dolphins leap out and catch the fish.[9] In lion (Panthera leo) pack hunting, each member of the hunting group is assigned a position, from left wing to right wing, in order to better obtain prey.[10] Such specializations in roles within the group are thought to increase sophistication in technique; lion wing members are faster, and will drive prey toward the center where the larger, stronger, killing members of the pride will take down the prey. Many observations of group pursuers note an optimal hunting size in which certain currencies (mass of prey killed or number of prey killed) are maximized with respect to costs (kilometers covered or injuries sustained).[11][12] Groups size is often dependent on aspects of the environment: number of prey, prey density, number of competitors, seasonal changes, etc.[13]
Birds
While birds are generally believed to be individual hunters, there are a few examples of birds that cooperate during pursuits. Harris's hawks (Parabuteo unicinctus) have two cooperative strategies for hunting. One strategy involves a group of hawks surrounding prey hidden under some form of cover, while another hawk attempts to penetrate the prey's cover. The penetration attempt flushes the prey out from its cover where it is swiftly killed by one of the surrounding hawks. The second strategy is less commonly used. It involves a "relay attack" in which a group of hawks, led by a "lead" hawk, engage in a long chase for prey. The "lead" hawk will dive in order to kill the prey. If the dive is unsuccessful, the role of the "lead" shifts to another hawk who will then dive in another attempt to kill the prey. During one observed relay attack, 20 dives and hence 20 lead switches were exhibited.[14]
Other avian group pursuer species
- Aplomado falcon (Falco femoralis)[15]
- Lanner falcon (Falco biarmicus)[15]
- Laggar falcon (Falco jugger)[15]
- Red-necked falcon (Falco chicquera)[15]
Invertebrates
_carrying_dead_termites_(13848808434).jpg)
_(6027759338).jpg)
As in vertebrates, there are many species of invertebrates which actively pursue prey in groups and exhibit task specialization, but while the vertebrates change their behavior based on their role in hunting, invertebrate task delegation is usually based on actual morphological differences. The vast majority of eusocial insects have castes within a population which tend to differ in size and have specialized structures for different tasks.[16] This differentiation is taken to the extreme in the groups isoptera and hymenoptera, or termites and ants, bees, and wasps respectively.
Termites are detritivores and therefore do not exhibit pursuit behaviors, however ants and wasps can be highly predatory and are known to pursue prey in groups. For example, termite-hunting ants of the genus Pachycondyla, also known as Matabele ants, form raiding parties consisting of ants of different castes, such as soldier ants and worker ants.[17] Soldier ants are much larger than worker ants, with more powerful mandibles and more robust exoskeletons, and so they make up the front lines of raiding parties and are responsible for killing prey. Workers usually butcher and carry off the killed prey, while supporting the soldiers. The raiding parties are highly mobile and move aggressively into the colonies of termites, often breaking through their outer defenses and entering their mounds. The ants do not completely empty the mound of termites, instead they only take a few, allowing the termites to recover their numbers so that the ants have a steady stream of prey.[17]
Asian giant hornets, Vespa mandarinia, form similar raiding parties to hunt their prey, which usually consists of honeybees.[18] The giant hornets group together and as a team can decimate an entire honeybee colony, especially those of non-native European honeybees. Alone, the hornets are subject to attack by the smaller bees, who swarm the hornet and vibrate their abdomens to generate heat, collectively cooking the hornet until it dies.[18] By hunting in groups, the hornets avoid this problem.
Individual Pursuers
Vertebrates

Mammals
While most big cat species are individual, ambush predators, Cheetahs (Acinonyx jubatus) are pursuit predators. Widely known as the fastest terrestrial animal, with speeds reaching 61–64 miles per hour, cheetahs take advantage of their speed during chases.[19][20] However, their speed and acceleration also have disadvantages, as both can only be sustained for short periods of time. Studies show that cheetahs can maintain maximum speed for a distance of approximately 500 yards.[21] Due to these limitations, cheetahs are often observed running at moderate speeds during chases. There are claims that the key to cheetahs' pursuit success may not be just their speed. Cheetahs are extremely agile, able to maneuver and change directions at very high speeds in very short amounts of time. This extensive maneuverability can make up for unsustainable high speed pursuit, as it allows cheetahs to quickly close the distance between prey without increasing their speed when prey change direction.[22]
Birds
The Painted redstart (Myioborus pictus) is one of the most well documented flush pursuers. When flies, prey for redstarts, are alerted of the presence of predators, they respond by fleeing. Redstarts take advantage of this anti-predator response by spreading and orienting their easily noticeable wings and tails, alerting the flies, but only when they are in a position where the flies' escape path intersects with the redstart's central field of vision. When prey's path are in this field of vision, the redstart's prey capture rate is at it maximum. Once the flies begin to flee, the redstart begins to chase. It has been proposed that redstarts exploit two aspects of the visual sensitivity of their prey: sensitivity to the location of the stimulus in the prey's visual field and sensitivity to the direction of stimulus environment.[23] The effectiveness of this pursuit can also be explained by "rare enemy effect", an evolutionary consequence of multi-species predator-prey interactions.[23]
Invertebrates

Dragonflies are skilled aerial pursuers; they have a 97% success rate for prey capture.[24] This success rate is a consequence of the "decision" of which prey to pursue based on initial conditions. Observations of several species of perching dragonflies show more pursuit initiations at larger starting distances for larger size prey species than for much smaller prey. Further evidence points to a potential bias towards larger prey, due to more substantial metabolic rewards. This bias is in spite of the fact that larger prey are typically faster and choosing them results in less successful pursuits. Dragonflies high success rate for prey capture may also be due to their "interception" foraging method. Unlike tracking foraging methods, in which predators focus on closing in on the current position of their prey, the interception method has the dragonfly seeking the position directly ahead of their prey as a way of surmising a prey's future location. Perching dragonflies (Libellulidae family), the largest family of dragonflies, have been observed "staking out" high density prey spots prior to pursuit.[1] There are no noticeable distinctions in prey capture efficiency between male and female dragonflies. Further, percher dragonflies are bound by their visual range. They are more likely to engage in pursuit when prey come within a subtended angle of around 1-2 degrees. Angles greater than this are outside of a dragonflies visual range.[1]
Evolutionary Basis of the Behavior
Evolution as a countermeasure
Current theory on the evolution of pursuit predation suggests that the behavior is an evolutionary countermeasure to prey adaptation. Prey animals vary in their likelihood to avoid predation, and it is predation failure that drives evolution of both prey and predator.[25] Predation failure rates vary wildly across the animal kingdom; raptorial birds can fail anywhere from 20% to 80% of the time in predation, while predatory mammals usually fail more than half the time.[25] Prey adaptation drives these low rates in three phases: the detection phase, the pursuit phase, and the resistance phase.[26] The pursuit phase drove the evolution of distinct behaviors for pursuit predation.
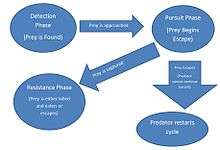
As selective pressure on prey is higher than on predators [25] adaptation usually occurs in prey long before the reciprocal adaptations in predators. Evidence in the fossil record supports this, with no evidence of modern pursuit predators until the late Tertiary period.[7] Certain adaptations, like long limbs in ungulates, that were thought to be adaptive for speed against predatory behavior have been found to predate predatory animals by over 20 million years. Because of this, modern pursuit predation is an adaptation that may have evolved separately and much later as a need for more energy in colder and more arid climates.[7] Longer limbs in predators, the key morphological adaptation required for lengthy pursuit of prey, is tied in the fossil record to the late Tertiary. It is now believed that modern pursuit predators like the wolf and lion evolved this behavior around this time period as a response to ungulates increasing feeding range.[7] As ungulate prey moved into a wider feeding range to discover food in response to changing climate, predators evolved the longer limbs and behavior necessary to pursue prey across larger ranges. In this respect, pursuit predation is not co-evolutionary with prey adaptation, but a direct response to prey. Prey's adaptation to climate is the key formative reason for evolving the behavior and morphological necessities of pursuit predation.
In addition to serving as a countermeasure to prey adaptation, pursuit predation has evolved in some species as an alternative, facultative mechanism for foraging. For example, polar bears typically act as specialized predators of seal pups and operate in a manner closely predicted by the optimal foraging theory. However, they have been seen to occasionally employ more energy-inefficient pursuit predation tactics on flightless geese. This alternative predatory strategy may serve as a back-up resource when optimal foraging is circumstantially impossible, or may even be a function of filling dietary needs.[27]
Evolution from an Ecological Basis
Pursuit predation revolves around a distinct movement interaction between predator and prey; as prey move to find new foraging areas, predators should move with them. Predators congregate in areas of high prey density,[28] and prey should therefore avoid these areas.[29] However, dilution factor may be a reason to stay in areas of high density due to a decreased risk of predation. Given the movements of predators over ranges in pursuit predation, though, dilution factor seems a less important cause for predation avoidance. Because of these interactions, spatial patterns of predators and prey are important in preserving population size. Attempts by prey to avoid predation and find food are coupled with predator attempts to hunt and compete with other predators. These interactions act to preserve populations.[30] Models of spatial patterns and synchrony of predator-prey relationships can be used as support for the evolution of pursuit predation as one mechanism to preserve these population mechanics. By pursuing prey over long distances, predators actually improve longterm survival of both their own population and prey population through population synchrony. Pursuit predation acts to even out population fluctuations by moving predatory animals from areas of high predator density to low predator density, and low prey density to high prey density. This keeps migratory populations in synchrony, which increases metapopulation persistence.[30] Pursuit predation’s effect on population persistence is more marked over larger travel ranges. Predator and prey levels are usually more synchronous in predation over larger ranges, as population densities have more ability to even out.[31] Pursuit predation can then be supported as an adaptive mechanism for not just individual feeding success but also metapopulation persistence.
Anti-predator Adaptation to Pursuit Predation
Anti-predator adaptation
Just as the evolutionary arms race has led to the development of pursuit behavior of predators, so too has it led to the anti-predator adaptations of prey. Alarm displays such as eastern swamphen's tail flicking, white-tailed deer's tail flagging, and Thomson's gazelles' stotting have been observed deterring pursuit.[32] These tactic are believed to signal that a predator's presence is known and, therefore, pursuit will be much more difficult. These displays are more frequent when predators are at an intermediate distance away. Alarm displays are used more often when prey believe predators are more prone to change their decision to pursue. For instance, cheetahs, common predators of Thomson's gazelles, are less likely to change their choice to pursue. As such, gazelles stott less when cheetahs are present than when other predators are present.[32] In addition to behavioral adaptations, there are also morphological anti-predator adaptations to pursuit predators. For example, many birds have evolved rump feathers that fall off with much less force than the feathers of their other body parts. This allows for easier escape from predator birds, as avian predators often approach prey from their rump.[33]
The Confusion Effect and Prey Oddity

In many species that fall prey to pursuit predation, gregariousness on a massive scale has evolved as a protective behavior and can be conspecific or heterospecific within a given group. This is primarily due to the Confusion Effect, which states that if prey animals congregate in large groups, predators will have more difficulty identifying and tracking specific individuals.[34] This effect has greater influence when individuals are visually similar and less distinguishable. In groups where individuals are visually similar, there is a negative correlation between group size and predator success rates. This may mean that the overall number of attacks decreases with larger group size or that the number of attacks per kill increases with larger group size.[35] This is especially true in open habitats, such as grasslands or open ocean ecosystems, where view of the prey group is unobstructed, in contrast to a forest or reef. Prey species in these open environments tend to be especially gregarious, with notable examples being starlings and sardines. When individuals of the herd are visually dissimilar, however, the success rate of predators increases dramatically.[35] In one study, wildebeest on the African Savannah were selected at random and had their horns painted white. This introduced a distinction, or oddity, into the population, and researchers found that wildebeest who had white horns were preyed upon at substantially higher rates.[36] By standing out, individuals are not as easily lost in the crowd, and so predators are able to track and pursue them with higher fidelity. This has been proposed as the reason why many schooling fish show little to no sexual dimorphism, and why many species in heterospecific schools bear a close resemblance to other species in their school.[34]
References
- 1 2 3 Combes, SA; Salcedo, MK; Pandit, MM; Iwasaki, JM (2013). "Capture Success and Efficiency of Dragonflies Pursuing Different Types of Prey". Integrative and Comparative Biology. 56 (5): 787–798. doi:10.1093/icb/ict072.
- ↑ Taylor, CR; Rowntree, VJ (1973). "Temperature regulation and heat balance in running cheetahs: A strategy for sprinters?". Am J Physiol. 224: 848–851.
- ↑ Creel, S.; Creel, N. M. (1995). "Communal hunting and pack size in African wild dogs, Lycaon pictus [Electronic version]". Animal Behaviour. 50 (5): 1325–1339. doi:10.1016/0003-3472(95)80048-4.
- ↑ Mech, D. (1970). The Wolf: The Ecology and Behaviour of an Endangered Species. Minneapolis, MN: University of Minnesota Press.
- ↑ Kruuk, H. (1972) The Spotted Hyena. University of Chicago Press, Chicago IL'
- ↑ DeVries, M.S.; Murphy, E.A.K.; Patek, S.N. (2012). "Strike mechanics of an ambush predator: the spearing mantis shrimp". Journal of Experimental Biology. 215: 4374–4384. doi:10.1242/jeb.075317.
- 1 2 3 4 5 Janis, C. M.; Wilhelm, P. B. (1993). "Were there mammalian pursuit predators in the Tertiary? Dances with wolf avatars". Journal of Mammalian Evolution. 1 (2): 103–125. doi:10.1007/bf01041590.
- ↑ Natural World Safaris. (2015, October 16). In 10 interesting facts about African wild dogs.
- ↑ Gazda, S. K.; Connor, R. C.; Edgar, R. K.; Cox, F. (2005). "A division of labour with role specialization in group-hunting bottlenose dolphins (Tursiops truncatus) off Cedar Key, Florida". Proceedings of The Royal Society. 272 (1559): 135–140. doi:10.1098/rspb.2004.2937.
- ↑ Stander, P.E. (1992). "Cooperative hunting in lions: the role of the individual". Behavioral Ecology and Sociobiology. 29: 445–454. doi:10.1007/bf00170175.
- ↑ Holekamp, K. E.; Smale, L.; Berg, R.; Cooper, S. M. (1996). "Hunting rates and hunting success in the spotted hyena (Crocuta crocuta) [Electronic version]". Journal of Zoology. 242 (1): 1–15. doi:10.1111/j.1469-7998.1997.tb02925.x.
- ↑ MacNulty, D. R.; Smith, D. W.; Mech, L. D.; Vucetich, J. A.; Packer, C. (2011). "Nonlinear effects of group size on the success of wolves hunting elk". Behavioral Ecology. 23 (1): 75–82. doi:10.1093/beheco/arr159.
- ↑ Packer, C.; Scheel, D.; Pusey, A. E. (1990). "Why lions form groups: food is not enough". American Naturalist. 136 (1): 1–19. doi:10.1086/285079.
- ↑ Bednarz, J. C. (1988). "Cooperative Hunting in Harris' Hawks (Parabuteo unicinctus)". Science. 239 (4847): 1525–1527. doi:10.1126/science.239.4847.1525.
- 1 2 3 4 Hector, D. P. (1986). "Cooperative Hunting and its Relationship to Foraging Success and Prey Size in an Avian Predator". Ethology. 73 (3): 247–257. doi:10.1111/j.1439-0310.1986.tb00915.x.
- ↑ Ross, Kenneth G.; Keller, Laurent (1995-01-01). "Ecology and Evolution of Social Organization: Insights from Fire Ants and Other Highly Eusocial Insects". Annual Review of Ecology and Systematics. 26: 631–656. doi:10.1146/annurev.ecolsys.26.1.631. JSTOR 2097222.
- 1 2 Leal, Inara R.; Oliveira, Paulo S. (1995-12-01). "Behavioral ecology of the neotropical termite-hunting ant Pachycondyla (= Termitopone) marginata: colony founding, group-raiding and migratory patterns". Behavioral Ecology and Sociobiology. 37 (6): 373–383. doi:10.1007/BF00170584. ISSN 0340-5443.
- 1 2 Sugahara, Michio; Sakamoto, Fumio (2009-06-24). "Heat and carbon dioxide generated by honeybees jointly act to kill hornets". Naturwissenschaften. 96 (9): 1133–1136. doi:10.1007/s00114-009-0575-0. ISSN 0028-1042. PMID 19551367.
- ↑ "10 Fastest Animals On Earth - Fastest Animals In The World." Conservation Institute. N.p., 06 Apr. 2014. Web. 16 Nov. 2015.
- ↑ Sharp, N. C. (2009). "Timed running speed of a cheetah". Journal of Zoology. 241 (3): 493–494. doi:10.1111/j.1469-7998.1997.tb04840.x.
- ↑ "How Fast Can Cheetah Run | Cheetah Built For Speed | Cheetah Serengeti." Barking Zebra Tours. N.p., 27 Sept. 2013. Web. 16 Nov. 2015.
- ↑ Wilson, A. M.; Lowe, J. C.; Roskilly, K.; Hudson, P. E.; Golabek, K. A.; Mcnutt, J. W. (2013). "Locomotion Dynamics of Hunting in Wild Cheetahs". Nature. 498 (7453): 185–89. doi:10.1038/nature12295. PMID 23765495.
- 1 2 Jablonski, P. G. "Sensory Exploitation of Prey: Manipulation of the Initial Direction of Prey Escapes by a Conspicuous 'rare Enemy'" Proceedings of the Royal Society B: Biological Sciences 268.1471 (2001): 1017-022. Web. 15 Oct. 2015
- ↑ Olberg RM, Worthington AH, Venator KR J Comp Physiol A. 2000 Feb; 186(2):155-62.
- 1 2 3 Geerat Vermeij,The American Naturalist Vol. 120, No. 6 (Dec., 1982), pp. 701-720
- ↑ Holling, C. S. (1966). The functional response of invertebrate predators to prey density. Memoirs of the Entomological Society of Canada, 98(S48), 5-86.
- ↑ Iles, D. T.; Peterson, S. L.; Gormezano, L. J.; Koons, D. N.; Rockwell, R. F. (2013). "Terrestrial predation by polar bears: Not just a wild goose chase". Polar Biology. 36 (9): 1373–1379. doi:10.1007/s00300-013-1341-5.
- ↑ Krebs, J. R. (1978). Optimal foraging: decision rules for predators. Behavioural ecology: an evolutionary approach, 23-63.
- ↑ Sih, Andrew (1984). "The Behavioral Response Race Between Predator and Prey". The American Naturalist. 123 (1): 143–150. doi:10.1086/284193.
- 1 2 Li, Z. Z.; Gao, M.; Hui, C.; Han, X. Z.; Shi, H. (2005). "Impact of predator pursuit and prey evasion on synchrony and spatial patterns in metapopulation". Ecological Modelling. 185 (2): 245–254. doi:10.1016/j.ecolmodel.2004.12.008.
- ↑ Rose, G. A.; Leggett, W. C. (1990). "The importance of scale to predator-prey spatial correlations: an example of Atlantic fishes". Ecology. 71: 33–43. doi:10.2307/1940245.
- 1 2 Hasson, Oren. "Pursuit-deterrent Signals: Communication between Prey and Predator." Trends in Ecology & Evolution 6.10 (1991): 325-29. Web. 9 Nov. 2015.
- ↑ Moller, A. P., J. T. Nielsen, and J. Erritzoe. "Losing the Last Feather: Feather Loss as an Antipredator Adaptation in Birds." Behavioral Ecology 17.6 (2006): 1046-056. Web.
- 1 2 Landeau, Laurie; Terborgh, John (1986). "Oddity And the 'Confusion Effect' in Predation.". Animal Behaviour. 34: 1372–1380. doi:10.1016/s0003-3472(86)80208-1.
- 1 2 Krakauer, David C. (1995). "Groups Confuse Predators by Exploiting Perceptual Bottlenecks: a Connectionist Model of the Confusion Effect.". Behavioral Ecology and Sociobiology. 36: 421–429. doi:10.1007/bf00177338.
- ↑ Kruuk, Hans (1973). "The Spotted Hyena. A Study Of Predation and Social Behavior.". The Journal of Animal Ecology. 42: 822. doi:10.2307/3145.