Peripheral nerve injury
Peripheral nerve damage is categorized in the Seddon classification based on the extent of damage to both the nerve and the surrounding connective tissue since the nervous system is characterized by the dependence of neurons on their supporting glia. Unlike in the central nervous system, regeneration in the peripheral nervous system is possible.[1] The processes that occur in peripheral regeneration can be divided into the following major events: Wallerian degeneration, axon regeneration/growth, and nerve reinnervation. The events that occur in peripheral regeneration occur with respect to the axis of the nerve injury. The proximal stump refers to the end of the injured neuron that is still attached to the neuron cell body; it is the part that regenerates. The distal stump refers to the end of the injured neuron that is still attached to the end of the axon; it is the part that will degenerate but remains the area that the regenerating axon grows toward. The study of peripheral nerve injury began during the American Civil War and has expanded to not only include extensive characterization of the processes and factors that contribute to nerve regeneration and reinnervation, but also determine therapies that enhance nerve regeneration such as biological conduits and the administration of growth-promoting molecules.[2]
Types of nerve damage
The lowest degree of nerve injury in which the nerve remains intact but its signaling ability is damaged is called neurapraxia. The second degree in which the axon is damaged, but the surrounding connecting tissue remains intact is called axonotmesis. The last degree in which both the axon and connective tissue are damaged is called neurotmesis.[2]
Overview of events in peripheral regeneration
Wallerian degeneration is a process that occurs before nerve regeneration and can be described as a cleaning or clearing process that essentially prepares the distal stump for reinnervation. Schwann cells are glial cells in the peripheral nervous system that support neurons by forming myelin that encases nerves. During Wallerian degeneration Schwann cells and macrophages interact to remove debris, specifically myelin and the damaged axon, from the distal injury site. (medscape) Calcium has a role in the degeneration of the damage axon. Bands of Büngner are formed when un-innervated Schwann cells proliferate and the remaining connective tissue basement membrane forms endoneurial tubes. Bands of Büngner are important for guiding the regrowing axon.[2]
At the neuronal cell body, a process called chromatolysis occurs in which the nucleus migrates to the periphery of the cell body and the endoplasmic reticulum breaks up and disperses. Nerve damage causes the metabolic function of the cell to change from that of producing molecules for synaptic transmission to that of producing molecules for growth and repair. These factors include GAP-43, tubulin and actin. Chromatolysis is reversed when the cell is prepared for axon regeneration.[3]
Axon regeneration is characterized by the formation of a growth cone. The growth cone has the ability to produce a protease that digests any material or debris that remains in its path of regeneration toward the distal site. The growth cone responds to molecules produced by Schwann cells such as laminin and fibronectin.[2]
Role of Schwann cells
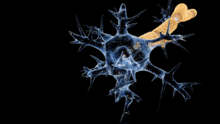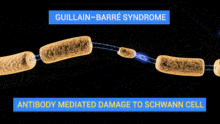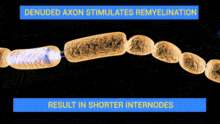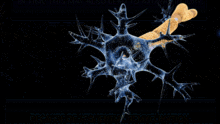
Schwann cells are active in Wallerian degeneration. They not only have a role in phagocytosis of myelin, but they also have a role in recruitment of macrophages to continue the phagocytosis of myelin. The phagocytic role of Schwann cells has been investigated by studying the expression of molecules in Schwann cells that are typically specific to inflammatory macrophages. Expression of one such molecule MAC-2, a galactose-specific lectin, is observed in not only degenerating nerves that are macrophage-rich but also degenerating nerves that are macrophage-scarce and Schwann cell-rich. Furthermore, the effects of MAC-2 in degenerating nerves are associated with myelin phagocytosis. There was a positive correlation between the amount of MAC-2 expression and the extent of myelin phagocytosis. A deficiency in MAC-2 expression can even cause inhibition of myelin removal from injury sites.[4]
Schwann cells are active in demylenation of injured nerves before macrophages are even present at the site of nerve injury. Electron microscopy and immunohistochemical staining analysis of teased nerve fibers shows that before macrophages arrive at the injury site, myelin is fragmented and myelin debris and lipid droplets are found in the cytoplasm of Schwann cells, indicating phagocytic activitiy before macrophages arrive.[5]
Schwann cell activity includes recruitment of macrophages to the injury site. Monocyte chemoattractant protein (MCP-1) plays a role in recruiting monocytes/macrophages. In tellurium-induced demylenation with no axon degeneration, nerve crush with axon degeneration, and nerve transection with axon degeneration an increase in MCP-1 mRNA expression followed by an increase in macrophage recruitment occurred. In addition varying levels of MCP-1 mRNA expression also had an effect. Increased MCP-1 mRNA levels correlated positively with an increase in macrophage recruitment. Furthermore, in situ hybridation determined that the cellular source of MCP-1 was Schwann cells.[6]
Schwann cells play an important role in not only producing neurotrophic factors such as nerve growth factor (NGF) and ciliary neurotrophic factor (CNTF), which promote growth, of both the damaged nerve and supporting Schwann cells, but also producing neurite promoting factors, which guide the growing axon, both of which are discussed below.
Role of macrophages
The primary role of macrophages in peripheral regeneration is demylenation during Wallerian degeneration. Immunohistochemical analysis showed that in tellurium demylenated, crushed, and cut nerves, expression of lysozyme, which is a marker for myelin phagocytosis, and of ED1, which is a marker for macrophages, occurred in the same region. Lysozyme was also investigated with respect to the temporal progression of myelin phagocytosis by macrophages in nerve injury. Northern blotting showed that peak lysozyme mRNA expression occurred at an appropriate time with respect to temporal models of myelin phagocytosis. Macrophages do not phagocytose all cellular debris at the nerve injury site; they are selective and will salvage certain factors. Macrophages produce apolipoprotien E which is involved in rescuing cholesterol in damaged nerves. In the same investigation, temporal levels of apolipoprotein E mRNA expression in the three models for demylenation and nerve damage were consistent with respect to models for cholesterol salvage in nerve injury. Macrophages play a role in salvaging cholesterol during nerve injury.[7]
Macrophages also play a role in inducing the proliferation of Schwann cells that occurs during Wallerian degeneration. Supernatant has been collected from medium in which macrophages are active in myelin phagocytosis where lysosomal processing of the myelin occurs within the macrophage. The supernatant contains a mitogenic factor, a mitosis promoting factor, that is characterized heat and trypsin sensitivity, both of which characterize it as a peptide. Treatment of Schwann cells with the collected supernatant shows that it is a mitogenic factor and thus plays an important role in the proliferation of Schwann cells.[8]
Macrophages are also involved in the secretion factors that promote nerve regeneration. Macrophages secrete not only interleukin-1, a cytokine that induces expression of nerve growth factor (NGF) in Schwann cells but also an interleukin-1 receptor antagonist (IL-1ra). Expression of IL-1ra in mice with transected sciatic nerves via implantation of a tube releasing IL-1ra showed the regrowth of fewer myelinated and unmyelinated axons. Macrophage secretion of interleukin-1 is involved in stimulation of nerve regeneration.[9]
Role of neurotrophic factors
Neurotrophic factors are those that promote survival and growth of neurons. A trophic factor can be described as a factor that is associated with providing nourishment to allow for growth. In general they are protein ligands for tyrosine kinase receptors; binding to the specific receptor yields autophosphorylation and subsequent phosphorylation of tyrosine residues on proteins that participate in further downstream signaling to acivate proteins and genes involved in growth and proliferation. Neurotrophic factors act through retrograde transport in neurons, in which they are taken up by the growth cone of the injured neuron and transported back to the cell body.[3][10]
Nerve growth factor (NGF) typically has a low level of expression in nerves that are healthy and not growing or developing, but in response to nerve injury NGF expression increases in Schwann cells. This is a mechanism to increase growth and proliferation of Schwann cells at the distal stump in order to prepare for reception of the regenerating axon. NGF has not only a trophic role but also a tropic or guiding role. The Schwann cells that form the bands of Bungner at the distal injury site express NGF receptors as a guiding factor for the regenerating axon of the injured neuron. NGF bound to the receptors on Schwann cells provides the growing neurons that are contacted with a trophic factor to promote further growth and regeneration[2][3][10]
Ciliary neurotrophic factor (CNTF) typically has a high level of expression in Schwann cells associated with nerves that are healthy, but in response to nerve injury CNTF expression decreases in Schwann cells distal to the injury site and remains relatively low unless the injured axon begins to regrow. CNTF has numerous trophic roles in motor neurons in the peripheral nervous system including the prevention of atrophy of dennervated tissue and the prevention of degeneration and death of motor neurons after nerve injury. (frostick) In sciatic motor neurons both CNTF receptor mRNA expression and CNTF receptor is increased after injury for a prolonged time frame compared to the short time frame in the central nervous system suggesting a role for CNTF in nerve regeneration.[11]
Insulin-like growth factors (IGFs) have been shown to increase the rate of peripheral nervous system axon regeneration. IGF-I and IGF-II mRNA levels are significantly increased distal to the site of crush injury in rat sciatic nerves.[12] At the site of nerve repair, locally delivered IGF-I can significantly increase the rate of axon regeneration within a nerve graft and help expedite functional recovery of a paralyzed muscle.[13][14]
Role of neurite-promoting factors
Neurite promoting factors include many extracellular matrix proteins produced by Schwann cells at the distal stump including fibronectin and laminin. Fibronectin are components of the basal lamina and promote neurite growth and adhesion of the growth cone to the basal lamina. In regenerating neural cells, neurite promoting factors play a role in adhesion of the axon and include neural cell adhesion molecule (N-CAM) and N-cadherin.[15]
Nerve regeneration therapies
Electrical stimulation can promote nerve regeneration. The frequency of stimulation is an important factor in the success of both quality and quantity of axon regeneration as well as growth of the surrounding myelin and blood vessels that support the axon. Histological analysis and measurement of regeneration showed that low frequency stimulation had a more successful outcome than high frequency stimulation on regeneration of damaged sciatic nerves.[16]
The use of autologous nerve grafting procedures that involve redirection of regenerative donor nerve fibers into the graft conduit has been successful in restoring target muscle function. Localized delivery of soluble neurotrophic factors may help promote the rate of axon regeneration observed within these graft conduits.[17]
An expanding area of nerve regeneration research deals with the development of scaffolding and bio-conduits. Scaffolding developed from biomaterial would be useful in nerve regeneration if they successfully exhibit essentially the same role as the endoneurial tubes and Schwann cell do in guiding regrowing axons.[18]
See also
- Nerve
- Neurapraxia
- Axonotmesis
- Neurotmesis
- Nerve injury
- Neuroregeneration
- Wallerian degeneration
- [[Peripheral nerve injury classif
ication]]
Notes
- ↑ Fenrich, Keith; Gordon, Tessa (2004). "Axonal Regeneration in the Peripheral and Central Nervous Systems - Current Issues and Advances". Canadian Journal of Neurological Sciences. 31 (2): 142–156. ISSN 0317-1671.
- 1 2 3 4 5 Campbell, William W. (31 August 2008). "Evaluation and management of peripheral nerve injury". Clinical Neurophysiology. 119 (9): 1951–1965. doi:10.1016/j.clinph.2008.03.018. PMID 18482862.
- 1 2 3 Burnett, Mark G.; Zager, Eric L. (2004). "Pathophysiology of Peripheral Nerve Injury: A Brief Review". Medscape Today: Neurosurgical Focus. ISSN 1092-0684. Retrieved 11 August 2013.
- ↑ Reichert, F; Saada, A; Rotshenker, S (May 1994). "Peripheral nerve injury induces Schwann cells to express two macrophage phenotypes: phagocytosis and the galactose-specific lectin MAC-2.". The Journal of neuroscience : the official journal of the Society for Neuroscience. 14 (5 Pt 2): 3231–45. PMID 8182468.
- ↑ Stoll, G.; Griffin, J. W.; Li, C. Y.; Trapp, B. D. (1 October 1989). "Wallerian degeneration in the peripheral nervous system: participation of both Schwann cells and macrophages in myelin degradation". Journal of Neurocytology. 18 (5): 671–683. doi:10.1007/BF01187086. PMID 2614485.
- ↑ Toews, Arrel D.; Barrett, Cheri; Morell, Pierre (15 July 1998). "Monocyte chemoattractant protein 1 is responsible for macrophage recruitment following injury to sciatic nerve". Journal of Neuroscience Research. 53 (2): 260–267. doi:10.1002/(SICI)1097-4547(19980715)53:2<260::AID-JNR15>3.0.CO;2-A. PMID 9671983.
- ↑ Venezie, R. D.; Toews, A. D.; Morell, Pierre (1 January 1995). "Macrophage recruitment in different models of nerve injury: Lysozyme as a marker for active phagocytosis". Journal of Neuroscience Research. 40 (1): 99–107. doi:10.1002/jnr.490400111. PMID 7714930.
- ↑ Baichwal,, R. R.; Bigbee,J. W.; Devries, G. H. (1988). "Macrophage-mediated myelin-related mitogenic factor for cultured Schwann cells.". Neurobiology. 85: 1701–1705. doi:10.1073/pnas.85.5.1701. JSTOR 31299. PMID 3422757.
- ↑ Guénard, V; Dinarello, CA; Weston, PJ; Aebischer, P (July 1991). "Peripheral nerve regeneration is impeded by interleukin-1 receptor antagonist released from a polymeric guidance channel.". Journal of neuroscience research. 29 (3): 396–400. doi:10.1002/jnr.490290315. PMID 1833560.
- 1 2 Frostick, Simon P.; Yin, Qi; Kemp, Graham J. (1 January 1998). "Schwann cells, neurotrophic factors, and peripheral nerve regeneration". Microsurgery. 18 (7): 397–405. doi:10.1002/(SICI)1098-2752(1998)18:7<397::AID-MICR2>3.0.CO;2-F. PMID 9880154.
- ↑ MacLennan, AJ; Devlin, BK; Neitzel, KL; McLaurin, DL; Anderson, KJ; Lee, N (1999). "Regulation of ciliary neurotrophic factor receptor alpha in sciatic motor neurons following axotomy.". Neuroscience. 91 (4): 1401–13. doi:10.1016/S0306-4522(98)00717-9. PMID 10391446.
- ↑ Glazner, Gordon W.; Morrison, Andrew E.; Ishii, Douglas N. (31 August 1994). "Elevated insulin-like growth factor (IGF) gene expression in sciatic nerves during IGF-supported nerve regeneration". Molecular Brain Research. 25 (3-4): 265–272. doi:10.1016/0169-328X(94)90162-7. PMID 7808226.
- ↑ Tiangco, David A.; Papakonstantinou, Konstantinos C.; Mullinax, Kelly A.; Terzis, Julia K. (1 January 2001). "IGF-I and End-to-Side Nerve Repair: A Dose-Response Study". Journal of Reconstructive Microsurgery. 17 (04): 247–256. doi:10.1055/s-2001-14516. PMID 11396586.
- ↑ Fansa, Hisham; Schneider, Wolfgang; Wolf, Gerald; Keilhoff, Gerburg (1 July 2002). "Influence of insulin-like growth factor-I (IGF-I) on nerve autografts and tissue-engineered nerve grafts". Muscle & Nerve. 26 (1): 87–93. doi:10.1002/mus.10165. PMID 12115953.
- ↑ Seckel, Brooke R. (1 September 1990). "Enhancement of peripheral nerve regeneration". Muscle & Nerve. 13 (9): 785–800. doi:10.1002/mus.880130904. PMID 2233865.
- ↑ Lu, M.-C.; Ho, C.-Y.; Hsu, S.-F.; Lee, H.-C.; Lin, J.-H.; Yao, C.-H.; Chen, Y.-S. (11 December 2007). "Effects of Electrical Stimulation at Different Frequencies on Regeneration of Transected Peripheral Nerve". Neurorehabilitation and Neural Repair. 22 (4): 367–373. doi:10.1177/1545968307313507. PMID 18663248.
- ↑ Thanos, PK; Okajima, S; Tiangco, DA; Terzis, JK (1999). "Insulin-like growth factor-I promotes nerve regeneration through a nerve graft in an experimental model of facial paralysis.". Restorative neurology and neuroscience. 15 (1): 57–71. PMID 12671244.
- ↑ Fansa, H; Keilhoff, G (March 2004). "Comparison of different biogenic matrices seeded with cultured Schwann cells for bridging peripheral nerve defects.". Neurological research. 26 (2): 167–73. doi:10.1179/016164104225013842. PMID 15072636.