Basilar membrane
| Basilar membrane | |
|---|---|
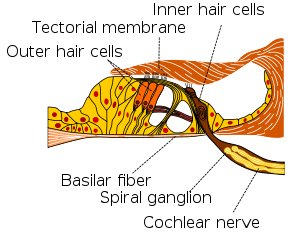 Section through organ of corti, showing basilar membrane | |
 Cross section of the cochlea. | |
| Details | |
| Identifiers | |
| Latin | membrana basilaris ductus cochlearis |
| MeSH | A09.246.631.246.125 |
The basilar membrane within the cochlea of the inner ear is a stiff structural element that separates two liquid-filled tubes that run along the coil of the cochlea, the scala media and the scala tympani (see figure).
Structure
The basilar membrane is a pseudo-resonant structure[1] that, like strings on an instrument, varies in width and stiffness. The "string" of the basilar membrane is not a set of parallel strings, as in a guitar, but a long structure that has different properties (width, stiffness, mass, damping, and the dimensions of the ducts that it couples to) at different points along its length. The motion of the basilar membrane is generally described as a traveling wave.[2] The parameters of the membrane at a given point along its length determine its characteristic frequency (CF), the frequency at which it is most sensitive to sound vibrations. The basilar membrane is widest (0.42–0.65 mm) and least stiff at the apex of the cochlea, and narrowest (0.08–0.16 mm) and most stiff at the base.[3] High-frequency sounds localize near the base of the cochlea (near the round and oval windows), while low-frequency sounds localize near the apex.
Function

Endolymph/perilymph separation
The fluids in these two tubes, the endolymph and the perilymph are very different chemically, biochemically, and electrically. Therefore, they are kept strictly separated. This separation is the main function of Reissner's membrane (between scala vestibuli and scala media), and it is also the function of tissue held by the basilar membrane such as the inner and outer sulcus cells (shown in yellow) and the reticular lamina of the organ of Corti (shown in magenta). For the organ of Corti the basilar membrane is permeable to perilymph. Here the border between endolymph and perilymph occurs at the reticular lamina, the endolymph side of the organ of Corti.[6]
A base for the sensory cells
The basilar membrane is also the base for the sensory cells of hearing, the hair cells that are equipped with "Stereocilia". There are approximately 15,000 hair cells in each human ear (see figure). This function as base of the sensory cells gave the basilar membrane its name, and it is again present in all land vertebrates. Due to its location, the basilar membrane places the hair cells in a position where they are adjacent to both the endolymph and the perilymph, which is a precondition of hair cell function.
Frequency dispersion
A third, evolutionarily younger, function of the basilar membrane is strongly developed in the cochlea of most mammalian species and weakly developed in some bird species:[7] the dispersion of incoming sound waves to separate frequencies spatially. In brief, the membrane is tapered and it is stiffer at one end than at the other. Furthermore, sound waves travelling to the far, "floppier" end of the basilar membrane have to travel through a longer fluid column than sound waves travelling to the nearer, stiffer end. Each part of the basilar membrane, together with the surrounding fluid, can therefore be thought of as a "mass-spring" system with different resonant properties: high stiffness and low mass, hence high resonant frequencies at the near end, and low stiffness and high mass, hence low resonant frequencies, at the far end.[8] This causes sound input of a certain frequency to vibrate some locations of the membrane more than other locations. As shown in experiments by Nobel Prize laureate Georg von Békésy, high frequencies lead to maximum vibrations at the basal end of the cochlear coil, where the membrane is narrow and stiff, and low frequencies lead to maximum vibrations at the apical end of the cochlear coil, where the membrane is wider and more compliant. This "place–frequency map" can be described quantitatively by the Greenwood function and its variants.
Sound-driven vibrations travel as waves along this membrane, along which, in humans, lie about 3,500 inner hair cells spaced in a single row. Each cell is attached to a tiny triangular frame. The 'hairs' are minute processes on the end of the cell, which are very sensitive to movement. When the vibration of the membrane rocks the triangular frames, the hairs on the cells are repeatedly displaced, and that produces streams of corresponding pulses in the nerve fibers, which are transmitted to the auditory pathway.[9] The outer hair cells feed back energy to amplify the traveling wave, by up to 65 dB at some locations.[10][11]
Additional images
-

Diagrammatic longitudinal section of the cochlea.
-

Floor of cochlear duct.
-
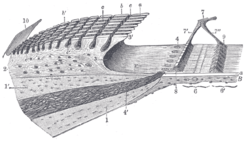
Spiral limbus and basilar membrane.
-
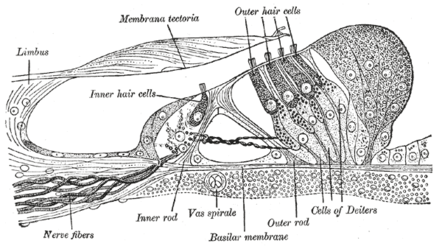
Section through the spiral organ of Corti (magnified)
-

The reticular membrane and subjacent structures.
See also
References
- ↑ M. Holmes and J. D. Cole, "Pseudoresonance in the cochlea, ' in: Mechanics of Hearing, E. de Boer and M. A. Viergever (editors), Proceedings of the IUTAM/ICA Symposium, Delft (1983), pp. 45-52.
- ↑ Richard R. Fay; Arthur N. Popper & Sid P. Bacon (2004). Compression: From Cochlea to Cochlear Implants. Springer. ISBN 0-387-00496-3.
- ↑ Oghalai JS. The cochlear amplifier: augmentation of the traveling wave within the inner ear. Current Opinion in Otolaryngology & Head & Neck Surgery. 12(5):431-8, 2004
- ↑ Shera, Christopher A. (2007). "Laser amplification with a twist: Traveling-wave propagation and gain functions from throughout the cochlea". Journal of the Acoustical Society of America. 122 (5): 2738–2758. doi:10.1121/1.2783205. Retrieved 13 April 2013.
- ↑ Robles, L.; Ruggero, M. A. (2001). "Mechanics of the mammalian cochlea". Physiological Reviews. 81 (3): 1305–1352. Retrieved 13 April 2013.
- ↑ Salt, A.N., Konishi, T., 1986. The cochlear fluids: Perilymph and endolymph. In: Altschuler, R.A., Hoffman, D.W., Bobbin, R.P. (Eds.), Neurobiology of Hearing: The Cochlea. Raven Press, New York, pp. 109-122
- ↑ Fritzsch B: The water-to-land transition: Evolution of the tetrapod basilar papilla; middle ear, and auditory nuclei. In: Douglas B. Webster; Richard R. Fay; Arthur N. Popper, eds. (1992). The Evolutionary biology of hearing. Berlin: Springer-Verlag. pp. 351–375. ISBN 0-387-97588-8.
- ↑ Schnupp J.; Nelken I.; King A. (2011). Auditory Neuroscience. Cambridge MA: MIT Press. ISBN 0-262-11318-X.
- ↑ Beament, James (2001). "How We Hear Music: the Relationship Between Music and the Hearing Mechanism". Woodbridge: Boydell Press: 97.
- ↑ Nilsen KE, Russell IJ (1999). "Timing of cochlear feedback: spatial and temporal representation of a tone across the basilar membrane". Nat. Neurosci. 2 (7): 642–8. doi:10.1038/10197. PMID 10404197.
- ↑ Nilsen KE, Russell IJ (2000). "The spatial and temporal representation of a tone on the guinea pig basilar membrane". Proc. Natl. Acad. Sci. U.S.A. 97 (22): 11751–8. doi:10.1073/pnas.97.22.11751. PMC 34345
 . PMID 11050205.
. PMID 11050205.
External links
- Auditory Neuroscience | The Ear several animations showing basilar membrane motion under various stimulus conditions
- Functional anatomy of the inner ear: plenty of images, animations, and very concise functional explanations
- Basilar Membrane Simulator Video and Scripts to Simulate the Basilar Membrane
- The role of the basilar membrane in sound reception: good explanation and diagrams